






Группа мелких карпофагов и карпонекрофагов в пустынях Средней Азии включает всех представителей родов Cardiocondyla, Pheidole, Monomorium и Tetramorium и Leptothorax из группы L. semenovi (L. semenovi, L. desertorum), т.е. почти всех мелких представителей подсемейства![]() Myrmicinae. Доля семян в питании этих муравьев может быть различной и в пустынях варьирует от 100% У Monomorium barbatulum до 50% У Pheidole arenarum. Питание последнего вида вообще весьма разнообразно. Он охотно посещает колонии тлей, собирает трупы насекомых и однажды мы даже наблюдали коллективную охоту Ph. arenarum на выплаживающихся в массе из разлагающегося детрита мелких мух. В Средней Азии он охотно поселяется в домах, где поедает буквально любые продукты. У Tetramorium schneideri семена составляют 80%, трупы насекомых — 15% и мелкие и малоподвижные живые насекомые — 5% добычи. Примерно такое же соотношение компонентов наблюдается и в питании T. striativentre.
Myrmicinae. Доля семян в питании этих муравьев может быть различной и в пустынях варьирует от 100% У Monomorium barbatulum до 50% У Pheidole arenarum. Питание последнего вида вообще весьма разнообразно. Он охотно посещает колонии тлей, собирает трупы насекомых и однажды мы даже наблюдали коллективную охоту Ph. arenarum на выплаживающихся в массе из разлагающегося детрита мелких мух. В Средней Азии он охотно поселяется в домах, где поедает буквально любые продукты. У Tetramorium schneideri семена составляют 80%, трупы насекомых — 15% и мелкие и малоподвижные живые насекомые — 5% добычи. Примерно такое же соотношение компонентов наблюдается и в питании T. striativentre.
И все же для всех представителей этой трофической группы семена являются наиболее важным и стабильным источником пищи, так что особенности использования ими кормового участка определяются в первую очередь именно карпофагией. В отличие от большинства зоонекрофагов мелкие карпофаги и карпонекрофаги производят мобилизацию из гнезд при нахождении обильного источника корма в любой точке кормового участка. В 1971 г. в Акыбае и в 1978 г. в Варганзе мы систематически выкладывали приманки из манной крупы на опытных площадках для выявления гнезд мелких![]() Myrmicinae. В процессе этой работы было зарегистрировано несколько сот актов групповой фуражировки у всех основных представителей группы. Эти данные сгруппированы в табл.28. Как видно из таблицы, максимальный радиус кормового участка (максимальное расстояние мобилизации) варьирует от 50 см у Cardiocondyla uljanini до 5 м у T. schneideri, а среднее расстояние, на котором производится мобилизация, — от 30 см у C. uljanini до 1,74 м у T. schneideri. Обнаруживается четкая положительная корреляция (r=0,897) между средним сухим весом рабочего (p) и средним расстоянием мобилизации (М), причем зависимость эта близка к прямолинейной и ближе всего описывается уравнением
Myrmicinae. В процессе этой работы было зарегистрировано несколько сот актов групповой фуражировки у всех основных представителей группы. Эти данные сгруппированы в табл.28. Как видно из таблицы, максимальный радиус кормового участка (максимальное расстояние мобилизации) варьирует от 50 см у Cardiocondyla uljanini до 5 м у T. schneideri, а среднее расстояние, на котором производится мобилизация, — от 30 см у C. uljanini до 1,74 м у T. schneideri. Обнаруживается четкая положительная корреляция (r=0,897) между средним сухим весом рабочего (p) и средним расстоянием мобилизации (М), причем зависимость эта близка к прямолинейной и ближе всего описывается уравнением
M=0,08+3,31p.
Явственно выражена и связь между размерами (весом) муравьев и максимальным расстоянием мобилизации, т.е. размером кормового участка. Связи между типом мобилизации и расстоянием, на которое она производится, обнаружить не удается. В то же время способ мобилизации сильно влияет на характер использования кормового участка и в первую очередь определяет скорость накопления муравьев в местах концентрации пищи.
Наиболее примитивную мобилизацию тандемом мы наблюдали только у Cardiocondyla uljanini, и у этого вида она в принципе не отличается от уже описанной мобилизации у североамериканских видов рода (Wilson, 1959). Разведчик, обнаруживший пищу, возвращаясь из гнезда к кормушке, может привести за собой одного нового рабочего. Этот рабочий в свою очередь может осуществить привод тандемом. Как и у изученных видов Cardiocondyla, у C. uljanini ведомый движется в непосредственной близости за лидером, постоянно прикасаясь к нему антеннами. Скорость накопления на кормушке при таком типе мобилизации невелика.
Наиболее распространенным способом мобилизации у мелких пустынных![]() Myrmicinae является мобилизация путем привода группы, сходная с описанной выше мобилизацией Camponotus. Мы наблюдали ее у всех видов Tetramorium, Monomorium barbatulum и Leptothorax semenovi. У всех этих муравьев разведчик, обнаруживший кормушку, возвращаясь в гнездо, оставляет кончиком брюшка пахучий след на субстрате. В гнезде он производит активацию и, если она оказывается успешной, выходит из гнезда в сопровождении группы активированных фуражиров. Все эти муравьи движутся к кормушке цепочкой, во главе которой обязательно находится разведчик. Иногда он при этом также метит тропу. След, оставленный во время предыдущего рейса, как правило, не используется и разведчик может привести группу к кормушке совсем не тем маршрутом, по которому он возвращался в гнездо. Как показали наши исследования на Myrmica, этот след служит для разведчика резервным ориентиром и используется при невозможности использования визуальных ориентиров (Длусский и др., 1978). Размер группы, идущей к кормушке, не превышает десяти особей и обычно цепочка состоит из трех — пяти муравьев. Возвращаясь в гнездо, рабочие, нашедшие с помощью разведчика кормушку, также обычно метят субстрат и по приходе в гнездо могут в свою очередь вывести на кормушку новые цепочки. Для мобилизации при помощи приводов характерно то, что настоящей тропы не образуется. Каждая группа и одиночные муравьи движутся своими собственными маршрутами. Однако, поскольку все они стремятся выбирать кратчайший путь, эти маршруты оказываются сходными, и поэтому поток муравьев, идущих к кормушке и к гнезду, внешне сходен с настоящей тропой. Характер накопления муравьев на кормушке при этом типе мобилизации был уже описан на примере Camponotus xerxes.
Myrmicinae является мобилизация путем привода группы, сходная с описанной выше мобилизацией Camponotus. Мы наблюдали ее у всех видов Tetramorium, Monomorium barbatulum и Leptothorax semenovi. У всех этих муравьев разведчик, обнаруживший кормушку, возвращаясь в гнездо, оставляет кончиком брюшка пахучий след на субстрате. В гнезде он производит активацию и, если она оказывается успешной, выходит из гнезда в сопровождении группы активированных фуражиров. Все эти муравьи движутся к кормушке цепочкой, во главе которой обязательно находится разведчик. Иногда он при этом также метит тропу. След, оставленный во время предыдущего рейса, как правило, не используется и разведчик может привести группу к кормушке совсем не тем маршрутом, по которому он возвращался в гнездо. Как показали наши исследования на Myrmica, этот след служит для разведчика резервным ориентиром и используется при невозможности использования визуальных ориентиров (Длусский и др., 1978). Размер группы, идущей к кормушке, не превышает десяти особей и обычно цепочка состоит из трех — пяти муравьев. Возвращаясь в гнездо, рабочие, нашедшие с помощью разведчика кормушку, также обычно метят субстрат и по приходе в гнездо могут в свою очередь вывести на кормушку новые цепочки. Для мобилизации при помощи приводов характерно то, что настоящей тропы не образуется. Каждая группа и одиночные муравьи движутся своими собственными маршрутами. Однако, поскольку все они стремятся выбирать кратчайший путь, эти маршруты оказываются сходными, и поэтому поток муравьев, идущих к кормушке и к гнезду, внешне сходен с настоящей тропой. Характер накопления муравьев на кормушке при этом типе мобилизации был уже описан на примере Camponotus xerxes.
Более эффективная система мобилизации наблюдается у ряда видов Monomorium, обитающих в аридных и субаридных районах Палеарктики: M. subopacum, M. venustum (Szlep, Jacobi, 1967), M. kusnezovi (Длусский и др., 1978) и ![]() M. gracillimum. Разведчик M. kusnezovi, обнаруживший корм, как и в предыдущем случае, оставляет на субстрате пахучий след кончиком брюшка. Если на пути к гнезду он встречает других рабочих, он контактирует с ними антеннами, после чего проконтактировавшие особи направляются по следу к кормушке. В гнезде разведчик производит направленную специфическую активацию пассивных фуражиров при помощи осцилляторной рекрутирующей демонстрации (ОРД) — характерных движений, пробежек, сочетающихся с антеннальными контактами. В процесс включаются также активаторы — рабочие, имевшие контакт с разведчиком, вернувшимся с кормушки. Они могут сами начать совершать ОРД. Антеннальные контакты играют в ОРД важнейшую роль. Прямые наблюдения в искусственных гнездах показали, что из гнезда выходят к кормушке только особи, имевшие тактильные контакты с разведчиком или активаторами.
M. gracillimum. Разведчик M. kusnezovi, обнаруживший корм, как и в предыдущем случае, оставляет на субстрате пахучий след кончиком брюшка. Если на пути к гнезду он встречает других рабочих, он контактирует с ними антеннами, после чего проконтактировавшие особи направляются по следу к кормушке. В гнезде разведчик производит направленную специфическую активацию пассивных фуражиров при помощи осцилляторной рекрутирующей демонстрации (ОРД) — характерных движений, пробежек, сочетающихся с антеннальными контактами. В процесс включаются также активаторы — рабочие, имевшие контакт с разведчиком, вернувшимся с кормушки. Они могут сами начать совершать ОРД. Антеннальные контакты играют в ОРД важнейшую роль. Прямые наблюдения в искусственных гнездах показали, что из гнезда выходят к кормушке только особи, имевшие тактильные контакты с разведчиком или активаторами.
В том случае, когда семья возбуждается слабо, картина мобилизации внешне сходна с таковой у Tetramorium: из гнезда выходят небольшие группы муравьев, во главе которых движется разведчик. Однако это сходство только внешнее. Группы всегда движутся по следу, оставленному разведчиком на пути от кормушки к гнезду, повторяя все его изгибы. Изъятие разведчика не приводит к нарушению мобилизации, и пассивные фуражиры все равно достигают кормушки по следу. В том случае, когда в гнезде имеется много пассивных фуражиров, готовых к мобилизации (например, если семья давно не получала пищи), они начинают выходить еще в то время, когда разведчик находится в гнезде, и самостоятельно по следу достигают кормушки. Для муравьев, не активированных ОРД и тактильными стимулами, след не является сигналом появления пищи на кормовом участке, и они пересекают его, не сворачивая к кормушке. Обычно разведчик, возвращаясь из гнезда к кормушке, повторно метит тропу. Метят ее и почти все рабочие, возвращающиеся в гнездо. Однако, поскольку муравьи не идут по следу совершенно точно, а могут отклоняться на расстояние до 1 см в сторону, постепенно изгибы трека первого фуражира сглаживаются и тропа спрямляется. Характер накопления особей на кормушке при этом типе мобилизации показан на рис:77.
Один из наиболее эффективных способов мобилизации наблюдается у Pheidole arenarum (Длусский, 1976). У этого вида имеются две четко обособленные касты — рабочие и солдаты. Активными фуражирами являются почти исключительно рабочие особи. После нахождения источника обильной пищи разведчик насыщается и движется к гнезду обычно по прямой, прикасаясь брюшком к субстрату и оставляя пахучий след. Рабочие, случайно обнаружившие свежий след, направляются по нему к кормушке без предварительного контакта с разведчиком. По пути к гнезду разведчик контактирует со всеми встреченными муравьями. Такие особи сразу возбуждаются, находят след и также приходят к кормушке. Войдя в гнездо, разведчик продолжает оставлять пахучий след, чего не наблюдается ни у одного из изученных нами муравьев. Одновременно он обменивается ударами антенны со встреченными муравьями. Обычно последние после контакта возбуждаются. Некоторые из них вскоре успокаиваются, другие переходят в предвходовую камеру, а часть покидают гнездо и по следу направляются к кормушке. Присутствие разведчика в этой группе не обязательно. Некоторые особи покидают гнездо без контакта с разведчиком или активаторами, возбужденные оставленным в гнезде следом. Многие мобилизованные муравьи также оставляют пахучий след по пути как к кормушке, так и к гнезду и таким образом формируют пахучую тропу. Через некоторое время количество муравьев на кормушке и интенсивность движения на тропе стабилизируются (рис:78/6/).
По мере насыщения семьи количество фуражирующих муравьев постепенно падает. Если во время активной групповой фуражировки убрать кормушку, муравьи долгое время продолжают ходить по тропе. Так, например, 3 мая 1970 г. в Акыбае в 1 м от гнезда Ф-701 была выставлена кормушка с сахарным сиропом в 9 ч 50 мин. В 9 ч 55 мин ее обнаружил первый разведчик и в 9 ч 59 мин началась массовая мобилизация. К 10 ч 15 мин количество муравьев на кормушке достигло 49 особей и в дальнейшем стабилизировалось и держалось в пределах 36—46 особей. В 12 ч 05 мин кормушка была убрана, но до 16 ч 20 мин муравьи продолжали ходить по тропе. До начала опыта контрольную линию на пути между гнездом и кормушкой в среднем пересекало 0,17 особей/мин, в первые 10 мин опыта — 0,7 особей/мин, в первые 10—20 мин после начала опыта — 5,2 особей/мин, в первые 20—30 мин после начала опыта — 11,9 особей/мин. В дальнейшем до конца опыта интенсивность движения сохранялась постоянной. В 15 ч, т.е. через 3 ч после того как убрали кормушку, интенсивность движения по тропе была 0,7 особей/мин, а к 16 ч в связи с общим увеличением активности муравьев (стало прохладнее) возросла до 4,2 особей/мин. В 16 ч 20 мин на этом гнезде был начат новый опыт. Следует отметить, что все это время тропа находилась в тени.
Наиболее интересное явление, обнаруженное нами у Ph. arenarum, — изменение соотношения рабочих и солдат при работе на разной пище. В описанном выше опыте на гнезде Ф-701 солдаты составляли в среднем 1,75±0,42% работающих особей (σ=1,04; n= 6). В тот же день в 16 ч 20 мин в 1 м от этого гнезда и от соседнего гнезда Ф-702 были выложены трупы крупных чернотелок. Опыт продолжался 3 ч 20 мин. Солдаты составляли в среднем 4,4±0,8% работающих особей в гнезде Ф-701 (σ=2,36, n=8) и 6,9+0,9% (σ=2,31; n=6) в гнезде Ф-702. Целесообразность увеличения числа солдат при доставке в гнездо трупа крупного насекомого очевидна, так как основной функцией этой подкасты является расчленение твердой пищи (трупов, семян).
Для более детального изучения этого процесса в 1974 г. на Звенигородской биостанции МГУ была проведена серия опытов с семьей Ph. arenarum, приведенной из Карши. Количество солдат в этой семье было специально взято выше, чем в норме (50% против 10% в норме). Семья, состоявшая из 14 самок, 141 рабочего, 139 солдат и большого количества расплода, была помещена в гипсовое гнездо, соединенное с ареной размером 14X62,5 см (рис:78/2/). Арена делилась стеклянной стенкой длиной 50 см на две равные субарены. Во время опытов на расстоянии 50 см от гнезда выставлялась кормушка и после начала мобилизации подсчитывалось количество муравьев на кормушке, интенсивность движения (количество муравьев, пересекающих контрольную линию на пути к кормушке за минуту) и относительное количество солдат. В опыте 1 (18 июля 1974 г.) на арену поместили кормушку с сахарным сиропом, в опыте 2 (19 июля) — труп крупного насекомого (бражника), в опыте 3 (21 июля) — кормушку с сиропом, в опыте 4 (22 июля) — 20, а затем еще 30 куколок Myrmica. На тропе относительное число солдат в первых трех опытах (в опыте 4 мобилизации не произошло) в первые 30 мин возрастает до 30—40%, а затем в течение часа постепенно падает до 10—13%. На кормушках с сахарным сиропом также вначале происходит увеличение, а к концу — уменьшение процента солдат. На трупе же относительное число солдат постоянно увеличивалось в течение опыта (рис:78/4/). На основании этих опытов можно предположить, что во всех случаях разведчики мобилизуют одинаковое число солдат, однако на сахарном сиропе насытившиеся солдаты перестают выходить из гнезда, а при нахождении трупа остаются на нем и продолжают работу. Это подтверждается и прямыми наблюдениями за мечеными особями. В опыте 4 на арене работали только рабочие. За все время наблюдений (2 ч 35 мин) произошла лишь одна небольшая (семь рабочих) мобилизация через 1 ч 20 мин после начала опыта. Один рабочий способен перенести в гнездо куколку, поэтому муравьи доставляли добычу в гнездо и там расчленяли.
Следует отметить, что неспецифическая активация у этого вида не наблюдается. В субарене без кормушки после мобилизации интенсивность движения падала в опыте 1 с 0,2 (контроль) до 0,07 особей/мин (опыт), в опыте 2 — с 0,4 до 0,06 особей/мин и в опыте 3 — с 0,07 до 0,02 особей/мин. В опыте 4 в контроле средняя интенсивность движения в субарену была 0,20 особей/мин, в опыте: в субарену с кормом — 0,22 особей/мин и в субарену без корма — 0,19 особей/мин. Таким образом, процесс организации групповой фуражировки у Ph. arenarum отличается рядом специализированных черт и во многом сходен с процессом, описанным у Solenopsis invicta (Wilson, 1962). Такими чертами являются отсутствие неспецифической активации, оставление разведчиком пахучего следа в гнезде и использование пахучего следа неактивированными рабочими.
Возникновение мобилизации с помощью пахучего следа в сочетании с ОРД на базе мобилизации за счет приводов, по-видимому, не представляет особой трудности. Во всяком случае в наших экспериментах с разными видами Myrmica, для которых в типе характерна именно мобилизация за счет приводов, нам удавалось довольно быстро приучить муравьев мобилизоваться по следу первого разведчика. И все же, несмотря на эффективность такой мобилизации, она совсем не характерна для истинно пустынных мелких карпофагов и карпонекрофагов, как,, например, различные Tetramorium или Monomorium barbatulum. Виды, использующие для мобилизации следовые феромоны, в настоящей пустыне встречаются довольно редко и больше характерны для влажных местообитаний — солончаков и тугаев (M. kusnezovi), оазисов (Ph. arenarum) или саксауловых лесов (![]() M. gracillimum). По-видимому, эта объясняется двумя причинами: во-первых, как уже отмечалось выше, на сухой горячей почве пахучий след плохо сохраняется; во-вторых, во время мобилизации при помощи следа вся масса пассивных фуражиров направляется в один участок и перестроить уже начавшуюся групповую фуражировку довольно сложно. В то же время мобилизация приводами позволяет семье одновременно организовать групповую фуражировку во многих точках кормового участка сравнительно небольшими группами особей, не закрывая при этом возможности направления всех пассивных фуражиров на одну кормушку. В условиях бедных кормом пустынных экосистем это дает несомненные преимущества. Так, например, у Ph. arenarum и T. striativentre и максимальное, и среднее расстояния мобилизации одинаковы (табл.28). При выкладывании приманок в Варганзе семьи T. striativentre делали до восьми дорог одновременно и эксплуатировали обычно все кормушки, находящиеся на кормовом участке. На тех же площадках у Ph. arenarum мы лишь в одном случае наблюдали три дороги, а обычно их было две или одна. В результате часть приманок на кормовом участке всегда оказывалась нетронутой.
M. gracillimum). По-видимому, эта объясняется двумя причинами: во-первых, как уже отмечалось выше, на сухой горячей почве пахучий след плохо сохраняется; во-вторых, во время мобилизации при помощи следа вся масса пассивных фуражиров направляется в один участок и перестроить уже начавшуюся групповую фуражировку довольно сложно. В то же время мобилизация приводами позволяет семье одновременно организовать групповую фуражировку во многих точках кормового участка сравнительно небольшими группами особей, не закрывая при этом возможности направления всех пассивных фуражиров на одну кормушку. В условиях бедных кормом пустынных экосистем это дает несомненные преимущества. Так, например, у Ph. arenarum и T. striativentre и максимальное, и среднее расстояния мобилизации одинаковы (табл.28). При выкладывании приманок в Варганзе семьи T. striativentre делали до восьми дорог одновременно и эксплуатировали обычно все кормушки, находящиеся на кормовом участке. На тех же площадках у Ph. arenarum мы лишь в одном случае наблюдали три дороги, а обычно их было две или одна. В результате часть приманок на кормовом участке всегда оказывалась нетронутой.
У всех представителей группы мелких карпофагов и карпонекрофагов кормовой участок не охраняется и участки соседних близко расположенных гнезд могут перекрываться. Охрана границы возникает лишь в том случае, когда сильно возрастает плотность муравьев, как это показал А. А. Захаров (1972), выкладывая приманки в зону, где работали особи из разных гнезд M. kusnezovi. Форма участка и его размеры во многом определяются характером гнездостроения. В том случае, когда семья обитает в односекционном гнезде и муравьи не строят системы тоннелей (например, T. striativentre или L. semenovi), форма кормового участка близка к круговой и его радиус равен длине максимального поискового рейда активных фуражиров (рис:79).
T. schneideri обычно строят многооекционные гнезда, секции которых располагаются неподалеку друг от друга. Население каждой секции представляет собой колонну, и перехода как активных, так и пассивных фуражиров из секции в секцию практически не происходит (Захаров, 1976). Фуражиры каждой секции используют только часть общего кормового участка семьи в пределах одного сектора (рис:80). Иногда между секторами наблюдаются даже нейтральные зоны, так же, как между территориями разных колонн у Formica. Активные фуражиры у этого вида имеют индивидуальные поисковые участки, а их относительное число (20% от общего числа фуражиров) необычайно велико для муравьев-карпофагов (Захаров, 1976). Все это вместе взятое позволяет T. schneideri создать систему использования территории, основанную на вторичном делении кормового участка, аналогичную системе Acantholepis semenovi или некоторых Formica. Но в то же время этот вид может осуществлять мобилизацию небольших групп пассивных фуражиров в участки концентрации пищи, что очень важно при сборе семян. По-видимому, такое сочетание вторичного деления территории и несовершенной мобилизации является оптимальным для мелкого пустынного карпонекрофага, и не случайно T. schneideri является единственным представителем своей размерной группы (p=0,25—1 мг) во всех типах пустынь на всей территории от Южного Казахстана до Афганистана. Лишь в Каршинской степи он местами заменяется сходным морфологически, но несколько более примитивным по поведению T. striativentre.
Система использования кормового участка у Ph. arenarum в некоторых отношениях сходна с системой T. schneideri. У этого вида также обычно имеются многосекционные гнезда, и население разных секций использует для фуражировки разные секторы кормового участка. Активные фуражиры ведут поиск на индивидуальных поисковых участках. У этого вида весьма развито взаимодействие особей на территории.
Активные фуражиры часто ходят в поиск парами: они то расходятся, то сходятся вновь и обмениваются ударами антенн. При обнаружении одним из них трупа насекомого второй помогает транспортировать добычу в гнездо. В то же время по ориентировочным оценкам активные фуражиры составляют у этого муравья около 8—10% от общего числа фуражиров, а мобилизация, как уже описывалась выше, чрезвычайно специализирована и оказывается целесообразной только при наличии долговременного и обильного источника пищи, что в настоящей пустыне наблюдается не так уж часто. Перейти же к менее совершенной мобилизации путем привода эти муравьи неспособны, так как для их рабочих запах следа является безусловным раздражителем, побуждающим их идти вдоль него в направлении от гнезда. В то же время в оазисах и особенно в жилищах человека эти особенности Ph. arenarum дают ему преимущества по сравнению с другими муравьями, имеющими сходные размеры.
У Monomorium kusnezovi и ![]() M. gracillimum характер использования территории довольно своеобразен. У этих муравьев имеется система поверхностных тоннелей, соединенных с главным гнездом. Вокруг каждого входа в тоннель фуражиры работают на небольшом расстоянии (обычно около 1 м), но общая площадь кормового участка может достигать 10 м2 и более. Как показали исследования А. А. Захарова (1972), активные фуражиры M. kusnezovi, составляющие 8% от общего числа фуражиров, имеют индивидуальные поисковые участки. С каждым тоннелем связана определенная группа пассивных фуражиров. Если запас пищи в районе входов в данный тоннель достаточно велик, они могут довольно долго находиться в этом тоннеле. Пассивные фуражиры могут у этого вида вести самостоятельный поиск добычи, но не производят мобилизации. Однако при изменении ситуации на кормовом участке происходит перераспределение фуражиров. Так, в одном из опытов А. А. Захарова (1972) в Акыбае в 1970 г. у гнезда с тремя тоннелями (рис:81) выставлялись кормушки с сахарным сиропом. Вначале кормушка была поставлена около тоннеля А. После мобилизации и двухчасовой групповой фуражировки кормушка была убрана. Однако в течение трех дней здесь находилось значительное число фуражиров. Через три дня несколько кормушек с сахарным сиропом и трупами насекомых были выставлены в районе тоннелей В и C. Этого оказалось достаточно, чтобы большинство пассивных фуражиров из тоннеля А перешло в тоннели В и С. Повторное выставление кормушек вблизи тоннеля А вызвало новое перераспределение фуражиров, однако большая часть муравьев, ранее работавших в этом тоннеле, осталась в тоннелях В и С. После того, как кормушки были убраны, пассивные фуражиры, промеченные в зонах различных тоннелей, вновь перераспределились. Таким образом вся система эксплуатации кормового участка у M. kusnezovi является более инертной, чем система Tetramorium, и более пригодна для участков, сравнительно богатых кормом. Вдобавок, поверхностные тоннели могут нормально функционировать только в достаточно влажной и плотной почве. Поэтому этот вид многочислен в оазисах и на солончаках и в настоящей пустыне встречается довольно редко, обычно на границе с оазисами.
M. gracillimum характер использования территории довольно своеобразен. У этих муравьев имеется система поверхностных тоннелей, соединенных с главным гнездом. Вокруг каждого входа в тоннель фуражиры работают на небольшом расстоянии (обычно около 1 м), но общая площадь кормового участка может достигать 10 м2 и более. Как показали исследования А. А. Захарова (1972), активные фуражиры M. kusnezovi, составляющие 8% от общего числа фуражиров, имеют индивидуальные поисковые участки. С каждым тоннелем связана определенная группа пассивных фуражиров. Если запас пищи в районе входов в данный тоннель достаточно велик, они могут довольно долго находиться в этом тоннеле. Пассивные фуражиры могут у этого вида вести самостоятельный поиск добычи, но не производят мобилизации. Однако при изменении ситуации на кормовом участке происходит перераспределение фуражиров. Так, в одном из опытов А. А. Захарова (1972) в Акыбае в 1970 г. у гнезда с тремя тоннелями (рис:81) выставлялись кормушки с сахарным сиропом. Вначале кормушка была поставлена около тоннеля А. После мобилизации и двухчасовой групповой фуражировки кормушка была убрана. Однако в течение трех дней здесь находилось значительное число фуражиров. Через три дня несколько кормушек с сахарным сиропом и трупами насекомых были выставлены в районе тоннелей В и C. Этого оказалось достаточно, чтобы большинство пассивных фуражиров из тоннеля А перешло в тоннели В и С. Повторное выставление кормушек вблизи тоннеля А вызвало новое перераспределение фуражиров, однако большая часть муравьев, ранее работавших в этом тоннеле, осталась в тоннелях В и С. После того, как кормушки были убраны, пассивные фуражиры, промеченные в зонах различных тоннелей, вновь перераспределились. Таким образом вся система эксплуатации кормового участка у M. kusnezovi является более инертной, чем система Tetramorium, и более пригодна для участков, сравнительно богатых кормом. Вдобавок, поверхностные тоннели могут нормально функционировать только в достаточно влажной и плотной почве. Поэтому этот вид многочислен в оазисах и на солончаках и в настоящей пустыне встречается довольно редко, обычно на границе с оазисами.
Наиболее специализированная и в то же время идеальная для карпофагии в участках с песчаной почвой и скудной растительностью система использования кормового участка наблюдается у Monomorium barbatulum. У этого вида имеются многосекционные гнезда, секции которого удалены друг от друга на расстояние около 1 м и соединены между собой горизонтальными галереями, проходящими на большой глубине и не разрушающимися во время сухого сезона. Население каждой секции обычно составляет 100—300 особей и эксплуатирует участок вокруг входа радиусом около 1—2 м. Территории разных секций перекрываются и таким образом вся семья использует для добывания пищи большую площадь. Помимо этого, во время влажного сезона функционируют системы поверхностных тоннелей более 1 м длиной с самостоятельными входами. Благодаря такой системе, сочетающейся с возможностью мобилизации приводами, этот вид может обитать даже на склонах барханов, почти лишенных растительности. Он является доминирующим мелким карпофагом во всех песчаных пустынях — от самых северных Астраханских песков до Афганистана. Однако в участках, более богатых пищей, он явно не выдерживает конкуренции с Monomorium kusnezovi, ![]() M. gracillimum и мелкими Tetramorium.
M. gracillimum и мелкими Tetramorium.
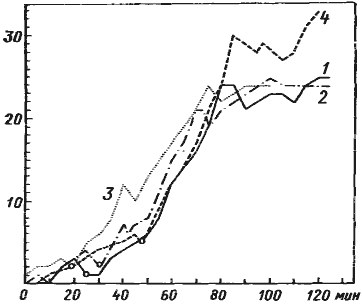
Рис. 77. Эффект мобилизации у Monomorium kusnezovi
(лабораторные эксперименты 1974 г.)
1 —3 — накопление фуражиров при нормальном режиме (кормление раз в сутки); 4 — накопление фуражиров после 2-суточного перерыва в кормлении. Все опыты поставлены на одной лабораторной семье. Кормушки с сахарным сиропом ставились в разных местах арены на расстоянии около 50 см от гнезда. За начало отсчета взят момент выставления кормушки на арену. Кружками обозначены моменты возвращения в гнездо первого фуражира с пищей. По вертикали — число особей на кормушке
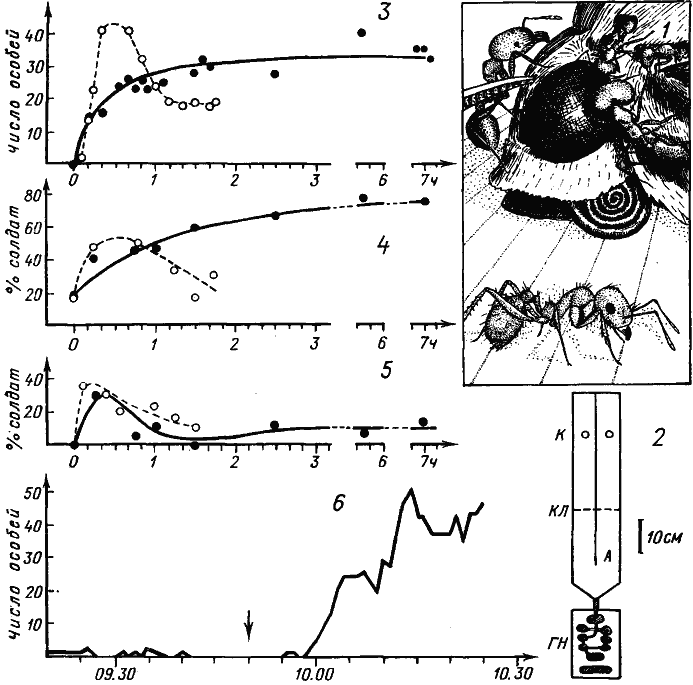
Рис. 78. Мобилизация и групповая фуражировка у Pheidole pallidula arenarum
1 — рабочие и солдаты Ph. arenarum на трупе бабочки. На переднем плане — рабочий, метящий субстрат. Расстояние между линиями на субстрате — 1 мм: 2 — чертеж установки, на которой проводились опыты: ГН — гнездо, А — арена, К — кормушки, КЛ — контрольная линия, на которой считались проходы муравьев; 3—5 — результаты двух лабораторных опытов с семьей, 50% особей которой составляли солдаты. На одну из субарен либо выставлялась кормушка с сахарным сиропом (пунктир, светлые кружки, опыт 18 июля 1974 г.), либо выкладывался труп крупного насекомого (сплошная линия, черные кружки, опыт 19 июля). По горизонтали — время с начала опыта: 3 — динамика числа особей на кормушке; 4 — доля солдат (в %) на кормушке; 5 — доля солдат (в %) среди особей, пересекавших контрольную линию; 6 — динамика накопления фуражиров на кормушке с сахарным сиропом в естественных условиях (Акыбай, 3 мая 1970 г., гнездо ФП-701). До 9 ч 50 мин учитывались муравьи, находившиеся в квадрате 20X20 см в метре от гнезда. В 9 ч 50 мин в центр этого квадрата поставлена кормушка. В дальнейшем муравьев считали только на кормушке (круг диаметром 5 см)
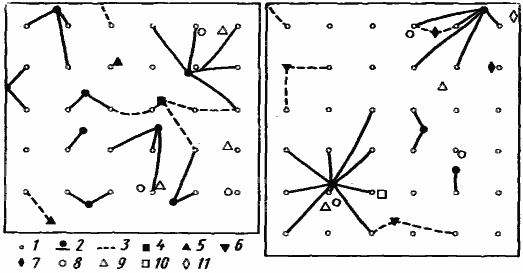
Рис. 79. Кормовые участки зерноядных муравьев на двух площадках в подгорной эфемеровой степи
(Варганза, май 1978 г.)
1 — кормушки с манной крупой; 2 — гнезда и дороги Tetramorium striativentre; 3 — дороги других зерноядных видов; 4 —7 — гнезда зерноядных муравьев: 4 — Messor aralocaspius, 5 — Monomorium kusnezovi, 6 — Pheidole arenarum, 7 — Tetramorium inerme; 8—11 — гнезда муравьев-зоонекрофагов: 8 — Plagiolepis sp., 9 — Proformica epinotalis, 10 — Acantholepis semenovi, 11 — Tapinoma karavaievi
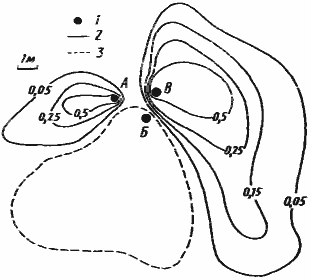
Рис. 80. Использование кормового участка трехсекционного гнезда Tetramorium schneideri при работе двух секций
[Захаров, 1976]
1 — входы в гнездо; 2 — изолинии динамической плотности на участках работающих секций (особей/ /дм2/мин); 3 — границы охраняемого участка секции Б, которая в день наблюдений не работала
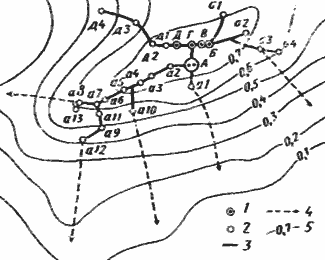
Рис. 81. Распределение особей на кормовом участке гнезда Monomorium kusnezovi
[Захаров, 1972]
1 — входы в кратеры; 2 — выходы из тоннелей; 3 — тоннели; 4 — основные направления фуражировки; 5 — изолинии равной динамической плотности (особей/дм2/мин) муравьев на участке
организм, питающийся семенами растений.
(зоонекрофаг), организм, питающийся мертвыми животными, гетеротроф
Всего 38 видов.
Всего насчитывает 549 видов.
Всего 415 видов
Всего 19 видов.
Старые названия: Acantholepis semenovi, Acantholepis transcaspiensis
Старое название: Leptothorax desertorum
Myrmicinae — самое большое подсемейство муравьев, включающее более 2 тыс. видов, относящихся более чем к 150 родам.
Старое название: Pheidole arenarum Ruzsky
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
участок, используемый муравьями для добычи пищи.
индивидуальное или групповое привлечение пассивных фуражиров активными фуражирами для конкретных полей в конкретном месте.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
Всего 965 видов.
Старое название: Leptothorax semenovi
галобионт
Subspecies: Monomorium indicum kusnezowi Santschi, 1928
 Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушитель
Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушительСтарое название: Monomorium gracillimum
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
у муравьев имеется три основные касты: самцы, самки и рабочие (бесплодные модифицированные самки).
(греч. φέρω — «нести» + ορμόνη — «гормон») — летучие вещества, управляющие поведенческими реакциями, процессами развития, а также многими процессами, связанными с социальным поведением и размножением. Феромоны модифицируют поведение, физиологическое и эмоциональное состояние или метаболизм других особей того же вида.
гнездо, состоящее из повторяющихся стереотипных блоков (секций), каждый из которых включает все видоспецифичные элементы (ствол, камеры и т. д.)
организационно оформленная мультифункциональная часть семьи (подсистема), имеющая постоянный состав и способная перейти к самостоятельному существованию.
часть гнезда рыжих лесных муравьев, занимаемая одной колонной.
пространство между охраняемыми территориями семей или колонн не посещаемое фуражирами из конкурирующих семей (колонн).
подразделение кормового участка семьи на элементарные поисковые участки, обслуживаемые одним фуражиром или небольшими группами
ксеробионт
Всего 60 видов.
гигрофил, галобионт
интенсивность посещения муравьями единицы площади в единицу времени (особь/дм²хмин.)
земляной вал при выходе из жилого муравейника (секции), не имеющего наземного купола.






