






Весьма разнородная группа мелких зоонекрофагов в пустынях Средней Азии представлена четырьмя видами: Plagiolepis pallescens (средний вес рабочего, р=0,08 мг), Acantholepis semenovi (р=0,23 мг), Tapinoma simrothi karavaievi (р=0,44 мг) и Crematogaster subdentata (р=0,51 мг). Ни один из этих муравьев не является настоящим специализированным пустынным видом. Первые два вида широко распространены в Средней Азии и в горах достигают лугово-степного пояса на высоте 1200—1400 м. Два других вида связаны главным образом с влажными местообитаниями, и в настоящей пустыне T. karavaievi обитает только на солончаках, a Cr. subdentata в саксауловых лесах. Ближайшие родственники всех этих муравьев обитают в семиаридных или гумидных областях. Численность семей достигает у них 10—100 тыс. особей, и уровень социальной организации довольно высок, особенно в сравнении с крупными дневными зоонекрофагами.
У P. pallescens гнездо обычно многосекционное, причем, поскольку вид олигогинен, некоторые секции содержат собственных самок. Питается он главным образом трупами насекомых и падью тлей. В некоторых типах пустынь весь кормовой участок семьи находится в пределах одного крупного куста полыни. Крупные трупы беспозвоночных муравьи расчленяют на месте и уносят в гнездо по частям. При этом всегда работают большие группы рабочих. Вообще, контакты на территории у этого муравья — обычное явление. Несколько раз удавалось наблюдать мобилизацию тандемом.
У Cr. subdentata, питающегося главным образом тлями и падью, одна семья занимает несколько изолированных гнезд, которые соединяются постоянными дорогами. Такие же постоянные дороги соединяют гнезда и кормовые почки (маленькие гнезда, не содержащие расплода) с колониями тлей. Внешне эта система напоминает систему ![]() Lasius fuliginosus, однако, как на самом деле происходит использование кормового участка у этого вида, сказать пока невозможно, так как экология его практически не изучена.
Lasius fuliginosus, однако, как на самом деле происходит использование кормового участка у этого вида, сказать пока невозможно, так как экология его практически не изучена.
У T. karavaievi, как и у Plagiolepis, имеются сложные многосекционные гнезда. Кормовые участки соседних гнезд обычно перекрываются, но при выкладывании приманок в зоне перекрытия появляется охраняемая граница (Захаров, 1972). У видов этого рода хорошо развита система мобилизации из гнезд, подробно описанная, в частности, у западного подвида T. simrothi phoenicea (Szlep, Jacobi, 1967). Муравей, обнаруживший кормушку, возвращается в гнездо, оставляя на пути пахучий след выделениями железы Павана на заднем крае 6-го брюшного стернита. Однако сам по себе этот след еще не является сигналом для других муравьев. Разведчик совершает в гнезде комплекс поведенческих действий, получивший название осцилляторной рекрутирующей демонстрации (ОРД. oscillatory recruitment display). ОРД состоит из пробежек по камерам, сочетающихся с антеннальными контактами и подергиваниями всем телом. В результате ОРД активируются пассивные фуражиры, которые выходят из гнезда и по следу идут к кормушке. Разведчик при этом может оставаться в гнезде. Поведение T. karavaievi — восточного подвида T. simrothi, по нашим наблюдениям, не отличается от описанного поведения T. phoenicea. Помимо мобилизации из гнезда, мы наблюдали у T. karavaievi также перемобилизацию с уже существующей тропы, которая может осуществляться двумя способами. В ряде случаев разведчик, обнаруживший новый источник корма, выходит на тропу и начинает совершать на ней ОРД. Собирается небольшая группа пассивных фуражиров. Затем эта группа вместе с разведчиком идет по новому следу, причем разведчик не просто идет впереди группы, как это наблюдается у Myrmica или Monomorium, а активно контактирует с другими особями (Захаров, 1972). Кроме того, как показали опыты С. И. Забелина (1979), рабочие T. karavaievi способны передавать информацию о нахождении корма на одном из ответвлений тропы при помощи тактильного кода. Экспериментальная установка представляла собой крест с лучами длиной около 15 см на подставке, стоявшей в ванночке с водой. Попасть на крест муравьи могли только по узкому мостику, ведшему в точку перекреста лучей. На двух из них находились кормушки с сахарным сиропом, прикрытые экранами из цветной бумаги. Муравьи, поднимавшиеся к установке по мостику, обычно контактировали антеннами со спускающимися фуражирами. Если контакт был только с одним из фуражиров, муравей поворачивал, как правило, к той кормушке, с которой возвращался этот фуражир. Этот механизм обеспечивает быстрое переключение части пассивных фуражиров на работу в новом участке (рис:72).
Различные способы организации групповой фуражировки сочетаются у T. karavaievi со способностью создавать долговременные дороги в участки, богатые пищей, и быстро строить в таких участках дополнительные гнезда, куда переселяется часть населения семьи вместе с расплодом. В результате у этого вида образуется своеобразная лабильная система использования кормового участка (Захаров, 1972). Несмотря на небольшие размеры муравьев, кормовые участки могут занимать площадь до 0,02 га. На этих участках имеется система дорог, по которым пассивные фуражиры ходят к колониям тлей на расстояние до 15 м от гнезда, а активные фуражиры выходят на индивидуальные поисковые участки не тратя времени на поиск в пути. Пища частично может доставляться в дополнительные гнезда, где находится часть расплода. При изменении ситуации вся эта система может за сравнительно короткое время перестраиваться, причем для образования новой дороги требуется не более часа, а для постройки нового гнезда один-два дня (рис:73). Следует, однако, подчеркнуть, что эта система не может нормально функционировать в условиях настоящей пустыни, где невозможно создать быстро дополнительное гнездо (почва либо плотная, либо слишком сухая на небольшой глубине) и пахучий след не может долго сохраняться на горячей поверхности почвы.
У A. semenovi кормовой участок используется совсем по иному принципу, что позволяет этому муравью обитать в самых различных стациях — от лугово-степи до почти полностью лишенных растительности пустынь. За исключением солончаков, где доминирует T. karavaievi, A. semenovi является единственным видом, занимающим трофическую нишу дневных зоонекрофагов 2-го размерного класса (р=0,1—0,25 мг) во всех типах пустынь Средней Азии. Лишь в северных пустынях Казахстана этот теплолюбивый вид сменяется Proformica epinotalis. Как показали исследования А. А. Захарова (1972), каждая семья A. semenovi имеет охраняемую территорию, на которую не допускаются муравьи из других семей того же вида. Между территориями разных семей имеются нейтральные зоны шириной 20—40 см (рис:74). Форма территории определяется характером распределения пищи и территориями соседних семей. В пределах ее можно выделить три зоны. В пригнездовой зоне с радиусом 20—30 см действуют в основном муравьи-строители, выносящие почву из гнезда. В ближней зоне охоты (до 2—3 м от гнезда) охота и транспортировка добычи ведутся коллективно. При появлении в этой зоне источника обильной пищи разведчики производят в гнездах неспецифическую активацию, приводящую к быстрому увеличению плотности муравьев. В дальней зоне охоты (до 8—10 м) активные фуражиры (сборщики трупов и одновременно охотники) ведут поиск на индивидуальных поисковых участках площадью около 0,25 м². Участки отдельных особей перекрываются. При обнаружении крупной добычи активация в гнезде не производится, но привлекаются другие особи, находящиеся в это время на территории. Рабочий, обнаруживший добычу, которую он не может самостоятельно унести в гнездо или даже умертвить, начинает возбужденно бегать вокруг нее, что привлекает муравьев, находящихся поблизости. В дальней зоне охоты работают и сборщики пади. К местам сбора пади они движутся по сходным траекториям, так что образуются потоки, которые, по мнению А. А. Захарова (1972), являются переходным этапом на пути к образованию постоянных дорог типа дорог Formica s. str. У внегнездовых рабочих этого вида, как и у Formica, четко выражен полиэтизм. Строители занимаются только переноской почвы или мусора, охотники собирают трупы или охотятся на мелких живых беспозвоночных и неохотно посещают кормушки с сахарным сиропом, трофобионты собирают падь тлей и не обращают внимания на приманки с белковой пищей (трупами насекомых). На территории постоянно можно наблюдать тактильные контакты между внегнездовыми рабочими.
Структура кормового участка, сходная с таковой у A. semenovi, вырабатывалась и у многих муравьев гумидной зоны (многие Formica), так что она не может рассматриваться как специальное приспособление, выработавшееся в связи с обитанием в пустынях. Однако преадаптивные возможности этой структуры в сочетании с особенностями гнездостроения (обитание в брошенных гнездах термитов, муравейниках крупных муравьев, в трещинах скал) позволили этому виду освоить зону пустынь. Род Acantholepis, и в частности группа A. frauenfeldi, куда относится A. semenovi, происходит из Африки. В Средней Азии эти муравьи — сравнительно недавние поселенцы. Весьма возможно, что прежде здесь существовали другие зоонекрофаги 2-го размерного класса (Proformica, мелкие Alloformica или мелкие Cataglyphis), вытесненные этим видом.
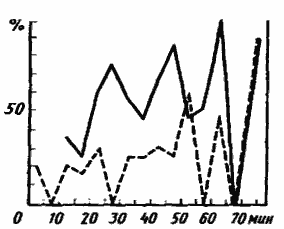
Рис. 72. Распределение доли проходов (в %) по новому следу у фуражиров Tapinoma karavaievi, имевших (сплошная линия) и не имевших (пунктир) тактильный контакт с возвращающимися фуражирами в последовательные 5-минутные интервалы [Забелин, 1970]
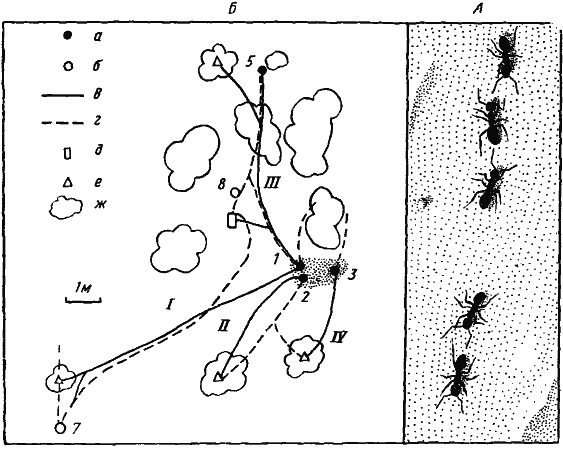
Рис. 73. Использование кормового участка у Tapinoma karavaievi
А — фуражиры, идущие по пахучему следу; Б — изменение системы дорог на кормовом участке после выставления кормушки (по [Захарову, 1972], изменено), а — входы в гнезда 30 апреля; б — входы, появившиеся к 6 мая 1970 г.; в — дороги 30 апреля; г — дороги 6 мая; д — кормушка с сахарным сиропом, выставленная 30 апреля и находившаяся на участке до 6 мая; е — колонии тлей; ж — кусты и деревья. Римскими цифрами обозначены номера дорог, арабскими — номера входов в гнезда
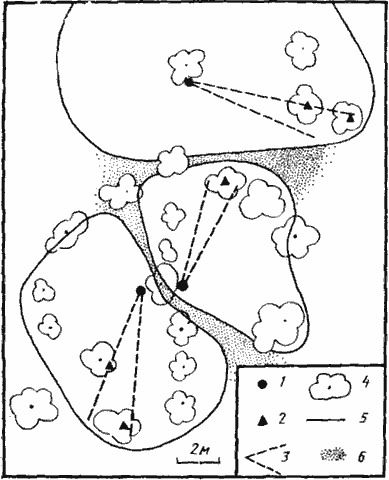
Рис. 74. План охраняемых территорий трех муравейников Acantholepis semenovi в Акыбае
[по Захарову, 1972]
1 — гнезда; 2 — колонии тлей; 3 — основные направления фуражировки; 4 — кусты; 5 — границы охраняемых территорий; 6 — нейтральные пространства
(зоонекрофаг), организм, питающийся мертвыми животными, гетеротроф
олигогинны
Старые названия: Acantholepis semenovi, Acantholepis transcaspiensis
гигрофил, галобионт
ситуация когда в гнезде живут несколько цариц, которые «не терпят» друг друга и занимают внутри муравейника отдельные территории. Смотри: моногиния, полигиния.
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
сладкая густая жидкость, выделяемая тлями, червецами и другими насекомыми, питающимися соком растений. Медоносная пчела собирает падь с листьев и перерабатывает в падевый мед.
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
участок, используемый муравьями для добычи пищи.
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
животные, не имеющие позвоночника: простейшие, губки, кишечнополостные, черви, моллюски, членистоногие, иглокожие и др.; примерно 3/4 видов всех животных.
индивидуальное или групповое привлечение пассивных фуражиров активными фуражирами для конкретных полей в конкретном месте.
 Lasius fuliginosus (Latreille, 1798) — пахучий муравей-древоточец, jet ants
Lasius fuliginosus (Latreille, 1798) — пахучий муравей-древоточец, jet ants(от греч. oikos дом, жилище, местопребывание и ...логия), наука об отношениях живых организмов и образуемых ими сообществ между собой и с окружающей средой. Термин «экология» предложен в 1866 Э. Геккелем. Объектами экологии могут быть популяции организмов, виды, сообщества, экосистемы и биосфера в целом.
Всего 60 видов.
гнездо, состоящее из повторяющихся стереотипных блоков (секций), каждый из которых включает все видоспецифичные элементы (ствол, камеры и т. д.)
сегмент брюшка.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
сигналы, передаваемые муравьями прикосновением антенн, щупиков, лапок при непосредственном контакте особей. Синоним: тактильный код.
участок, охраняемый семьей (колонной) от посещения чужими особями своего или близких видов.
пространство между охраняемыми территориями семей или колонн не посещаемое фуражирами из конкурирующих семей (колонн).
(от трофо… и греч. allaxis — обмен) передача пищи и гормональных веществ от особи к особи путем кормления содержимым зобика, желудка или слизывания выпота с поверхности тела. Играет огромную роль в передаче информации между муравьями.
выполнение муравьем в течение длительного времени определенных функций в семье. Если функция связана только с возрастом - возрастной полиэтизм; если функция выполняется постоянно - постоянный, или «кастовый», полиэтизм.
взаимовыгодные отношения между муравьями и выделяющими падь насекомыми: слизывая падь, муравьи охраняют этих насекомых.
отряд общественных насекомых. Общины, разделенные на касты, состоят из крылатых и бескрылых особей. Строят подземные и наземные (до 15 м высотой) гнезда (термитники). Ок. 2600 видов, главным образом в тропиках; в России 2 вида: один в районе Сочи, второй, вероятно, завезенный из Японии или Китая, во Владивостоке. Многоядны. Разрушают древесину, портят бумагу, кожу, сельскохозяйственные продукты и др. Термиты, обитающие в почве, полезны как почвообразователи.
Старое название: Acantholepis frauenfeldi
Всего 24 вида.
Всего 4 вида.
Всего 65 видов.






