






/Материал к этому разделу был собран
и подготовлен студенткой МГУ Е. Б. Федосеевой.
Ею также выполнены рисунки 18—20, 24./
Хорошо известна необычайно высокая подвижность многих дневных муравьев, обитающих в пустынях. Действительно, ни один из муравьев гумидных областей Палеарктики не может двигаться с такой высокой скоростью, как пустынные Cataglyphis. Очень быстро бегают также Proformica и Acantholepis и, судя по описаниям, Ocymyrmex и Anoplolepis Южной Африки, Melophorus Австралии, Dorymyrmex и Forelius Южной Америки и некоторые Myrmecocystus Северной Америки.
Во многих случаях увеличение скорости движения скоррелировано с увеличением длины конечностей. Как видно из табл.13, у Cataglyphis относительная длина конечностей явственно больше, чем у других представителей трибы Formicini, обитающих в семиаридных (Alloformica, Proformica) и гумидных (Formica) условиях. Исключение представляет лишь C. emeryi, наиболее примитивный представитель рода, поведение которого, как отмечалось выше, сходно с поведением Alloformica. Понятно, что увеличение длины конечностей обеспечивает большую ширину шага и соответственно большую скорость движения насекомого при той же частоте шагов. Из правил работы рычажной системы следует, что увеличение одного плеча рычага (конечности) требует соответственного увеличения действующей силы (мышцы) другого плеча. Вследствие этого увеличение длины конечности должно сопровождаться и увеличением относительной длины мышц, приводящих их в движение. Поскольку часть мышц, управляющих движением ног, располагается в груди, форма груди также будет изменяться. Удлинение мышц происходит, с одной стороны, за счет общего удлинения груди, а с другой — за счет смещения элементов внутреннего скелета, к которым крепятся эти мышцы.
Как это происходит, можно представить, сопоставив мускулатуру и внутренний скелет Alloformica aberrans и Cataglyphis setipes (рис:18, рис:19, рис:20). Как у одного, так и у другого вида движением задних ног управляют четыре мышцы, расположенные в проподеуме: три, связанные с коксой и одна — с вертлугом. Кокса сочленяется с плейритом в одной точке: отверстие, соединяющее полость коксы с полостью тела, несет сочленовный отросток, окруженный ободком. В основании ободка имеется впадина, в которую входит сочленовный отросток проподеума. M. protractor coxae /Названия мышц даны по работе Н. В. Насонова (1889). Реальная функция мышц иногда не соответствует названию./, отводящий коксу вперед, направляется от верхнейи боковой поверхности проподеума и мезопроподеальной фрагмы к наружному краю коксального ободка. M. flexor coxae, отводящий коксу назад, отходит от ветви задней фурки, диафрагмы и стернальной фрагмы проподеума и прикрепляется к внутреннему краю коксального ободка. Действие m. extensor coxae аналогично действию m. protractor coxae. Эта мышца крепится к диафрагме и стернальной фрагме проподеума, как и m. flexor coxae, но ее волокна отходят ниже и прикрепляются к внешнему краю коксального ободка рядом с m. protractor coxae. Разгибатель вертлуга (m. extensor trochanteris) отходит от переднего края проподеума в области мезопроподеального шва и от наружной части переднего конца ветви задней фурки и, пройдя через отверстие коксы, крепится к внутреннему краю вертлуга. Как видно из сказанного, мышцы, управляющие движением задних ног, крепятся к основанию проподеума, мезопроподеальной фрагме и задней фурке проподеума. Для увеличения длины мышц необходимо увеличить расстояние между коксой и этими элементами эндоскелета. Такое увеличение и наблюдается у Cataglyphis. Проподеум у этих муравьев (за исключением C. emeryi) относительно ниже и длиннее, чем у Alloformica, причем заметно удлиняется основная часть, что сопровождается вытягиванием ветвей задней фурки, связанной с основанием проподеума. У A. aberrans отношение длины ветвей к длине основания фурки составляет около 1.6, а у C. setipes — 3.0.
В результате этой перестройки координации меняется весь облик муравья: увеличение длины конечностей сопровождается удлинением груди и ее сужением в средней части. Интересно отметить, что у Cataglyphis, предки которых имели сравнительно длинный проподеум, сужается задняя часть среднегруди, а у Myrmecocystus и Prenolepis, предки которых (скорее всего, Lasius) имели короткий и высокий проподеум, перетяжка находится в средней части среднегруди (рис:21).
Описанное изменение пропорций связано не непосредственно с увеличением скорости движения, а с увеличением длины конечностей и ширины шага. Поэтому с подвижными пустынными муравьями обнаруживают конвергентное сходство некоторые муравьи тропических лесов, живущие в кронах деревьев (Oecophylla, некоторые Dolichoderus, Camponotus, Aphaenogaster; рис:22). Этим муравьям длинные конечности необходимы для того, чтобы перешагивать (а иногда и перепрыгивать) с листа на лист, не спускаясь на землю или на крупные ветви. Правда, у этих муравьев, в отличие от пустынных, одновременно удлиняется и голова, так как муравей, прежде чем перешагнуть с листа на лист, должен ощупать или обнюхать кончиками антенн дорогу впереди. К затылочным углам головы прикрепляются жевательные мышцы. Поэтому в тех случаях, когда удлинение головы не сопровождается увеличением длины жвал (и, следовательно, жевательных мышц), она приобретает своеобразную колбовидную форму (рис:22).
Также /известно/, что многие дневные виды во время движения в жаркую погоду держат брюшко вертикально вверх. Такое поведение независимо возникает у пустынных представителей подсемейств![]() Myrmicinae (Myrmicaria),
Myrmicinae (Myrmicaria), ![]() Dolichoderinae (Dorymyrmex, Forelius) и
Dolichoderinae (Dorymyrmex, Forelius) и![]() Formicinae (Cataglyphis). Широко распространено мнение, что поднятое вертикально вверх брюшко меньше нагревается, так как, во-первых, обращено к солнцу меньшей поверхностью, во-вторых, выше поднято над нагретой почвой и, в-третьих, на бегу лучше обдувается ветром. Кроме того, высказывалось предположение, что другой функцией этого поведения является оставление пахучей дорожки в воздухе, так как на подвижном песке след плохо сохраняется (Goetsch, 1953). Абсурдность последнего утверждения очевидна, так как в пустыне с постоянно дующими ветрами, конвекционными потоками от нагретой почвы и редким травяным покровом пахучий след в воздухе будет сохраняться еще хуже, чем на подвижном песке. Не менее наивно выглядит и утверждение о том, что обдувание брюшка ветром может его охлаждать. Ведь у муравьев нет потовых желез, как у человека, и испарение воды у них идет главным образом через дыхальца проподеума.
Formicinae (Cataglyphis). Широко распространено мнение, что поднятое вертикально вверх брюшко меньше нагревается, так как, во-первых, обращено к солнцу меньшей поверхностью, во-вторых, выше поднято над нагретой почвой и, в-третьих, на бегу лучше обдувается ветром. Кроме того, высказывалось предположение, что другой функцией этого поведения является оставление пахучей дорожки в воздухе, так как на подвижном песке след плохо сохраняется (Goetsch, 1953). Абсурдность последнего утверждения очевидна, так как в пустыне с постоянно дующими ветрами, конвекционными потоками от нагретой почвы и редким травяным покровом пахучий след в воздухе будет сохраняться еще хуже, чем на подвижном песке. Не менее наивно выглядит и утверждение о том, что обдувание брюшка ветром может его охлаждать. Ведь у муравьев нет потовых желез, как у человека, и испарение воды у них идет главным образом через дыхальца проподеума.
Даже у крупных особей C. setipes при подъеме брюшка его центр поднимается всего на 2—3 мм. Прямые измерения в Варганзе показали, что при температуре воздуха 29—30° и температуре на поверхности почвы 53—55° разница температур на высоте 5 и 10 мм от поверхности почвы составляет всего 1,5° (т.е. 0,3 градуса/мм). Трудно предположить, что столь незначительное преимущество могло привести к столь серьезной перестройке координации (изменение положения и длины мышц проподеума и петиолюса, формы петиолюса, смещение осей вращения). Аналогичный эффект легко мог бы быть достигнут изменением окраски брюшка, не затрагивающим сложившихся координаций.
Как нам кажется, гораздо ближе к истине К. В. Арнольди (1964), высказавший предположение, что задирание брюшка у Cataglyphis приводит к перемещению центра тяжести и каким-то образом связано с высокой скоростью движения этих муравьев. Почему-то до сих пор никто не обратил внимания на бросающийся в глаза факт, что подобное поведение вообще характерно для![]() Formicinae и
Formicinae и ![]() Dolichoderinae с длинными ногами, вне зависимости от того, в каком климате, аридном или гумидном, они обитают. Брюшко поднимают вверх Oecophylla, Leptomyrmex и некоторые другие муравьи, живущие в лесах, в том числе и влажных тропических (рис:22).
Dolichoderinae с длинными ногами, вне зависимости от того, в каком климате, аридном или гумидном, они обитают. Брюшко поднимают вверх Oecophylla, Leptomyrmex и некоторые другие муравьи, живущие в лесах, в том числе и влажных тропических (рис:22).
Для определения центра тяжести C. setipes при различных положениях брюшка была изготовлена увеличенная модель муравья в профиль из плотной бумаги. Для того чтобы соотношение весов головы, груди, петиолюса и брюшка было таким же, как у живых муравьев, к разным частям модели подклеивались дополнительные слои бумаги. Для определения центра тяжести модели вначале части ее жестко фиксировались в определенном положении. Затем модель свободно подвешивалась на игле за «брюшко» и от точки вращения проводилась при помощи отвеса отвесная линия. Затем модель подвешивалась за «голову» и проделывалась та же операция. Точка пересечения отвесных линий и принималась за центр тяжести. На рис:23 видно, что при подъеме брюшка центр тяжести перемещается криволинейно и смещается вперед.
Для того чтобы понять, какие преимущества дает изменение положения центра тяжести, необходимо рассмотреть, какие силы действуют на тело во время перемещения. Количество энергии, расходуемой мышцами конечностей, существенным образом определяется взаимодействием сил сокращения мускулов и силы тяжести, действующей на тело. При ходьбе и беге разные пары ног муравьев, как и большинства других насекомых, выполняют различные функции (Manton, 1952).
Толчок осуществляется главным образом задними ногами. Передние ноги подтягивают тело, а средние дают дополнительную точку опоры при переносе тела. Поэтому для простоты изложения мы можем рассматривать в качестве силы толчка результирующую сил сокращения мышц только задней конечности, а в качестве точки вращения — коленное сочленение передней конечности. Подобные упрощения не изменяют сути дела, но облегчают понимание. На схеме на рис:23 изображены векторы сил, действующих на тело муравья во время толчка. Ось вращения и векторы моментов сил направлены перпендикулярно чертежу; остальные векторы лежат в плоскости чертежа.
Введем следующие обозначения: О — ось вращения; А — центр тяжести тела; Р — сила тяжести; rP — радиус-вектор Р, соединяющий центр тяжести тела с осью вращения; β — угол между Р и rP; F — сила толчка задней ноги; rF — радиус-вектор F, соединяющий точку приложения этой силы (В) с осью вращения; α — угол между F и rF. Величина момента любой силы зависит от расстояния между точкой приложения силы и осью вращения (радиус-вектора), величины действующей силы и угла между направлением ее действия и радиус-вектором. Таким образом, величина момента силы толчка (MF) будет равна:
MF≈rF*sinα*F.
Но толчку будет противодействовать сила тяжести с моментом MP=rP*sinβ*P.
При перемещении центра тяжести вперед л вверх уменьшаются угол β и rP, а следовательно, и величина момента силы тяжести при постоянной Р. Это повышает эффективность толчка, т.е. в конечном счете дает увеличение скорости при тех же энергетических затратах.
Если это так, возникает законный вопрос: почему все муравьи-герпетобионты не поднимают брюшка во время бега? Для того чтобы разобраться в этом, необходимо вначале рассмотреть, как осуществляется подъем брюшка. Обычно при движениях брюшка происходит поворот в двух сочленениях — между проподеумом и петиолюсом и между петиолюсом и брюшком. Этими движениями управляют две группы мышц, расположенные соответственно в проподеуме и петиолюсе (рис:19, рис:20, рис:24). Петиолюс соединяется с задним краем проподеума при помощи сочленения, состоящего из отростка на верху стебелька и небольшого отверстия в проподеуме, ограниченного двумя отростками основного отверстия, через которое полость груди соединяется с полостью петиолюса. Движением петиолюса управляют четыре пары мышц, расположенных в проподеуме. Верхние мышцы, поднимающие петиолюс (m. elevator petioli superior), отходят от основной поверхности проподеума и прикрепляются к сочленовному отростку стебелька. M. depressor petioli, опускающие петиолюс, идут от внутренней поверхности ветвей задней фурки и крепятся к нижнему краю стебелька. Некоторые волокна этой мышцы могут отходить от мезопроподеальной фрагмы, образованной швом между тергитом среднегруди и проподеумом. M. elevator petioli inferior начинаются на основной поверхности проподеума и крепятся на боковых поверхностях переднего края стебелька. M. protractor petioli отходят от основания задней фурки проподеума и крепятся к боковым поверхностям переднего края стебелька выше m. elevator petioli inferior, образуя с ними перекрест. Эти две пары при совместном действии фиксируют положение петиолюса, а действуя поочередно поворачивают его вправо или влево. Кроме того, m. elevator petioli inferior могут участвовать в опускании стебелька, усиливая действия m. depressor petioli.
В поднимании и опускании брюшка принимают участие и две пары мышц, расположенных в петиолюсе. Петиолюс причленяется к брюшку при помощи шарнирного сочленения, образованного двумя впадинами в задней части стебелька и соответствующими отростками на ободке первого брюшного тергита, окружающем отверстие, соединяющего полости стебелька и брюшка. Опускающие брюшко m. flexor abdominalis у A. aberrans отходят от верхней части передней поверхности чешуйки и прикрепляются к нижнему краю отверстия первого сегмента брюшка. M. elevator abdominalis, поднимающие брюшко, отходят от нижней части передней поверхности чешуйки и прикрепляются к верхнему краю отверстия первого брюшного сегмента.
Если сравнить действие мышц, поднимающих и опускающих петиолюс у A. aberrans (рис:19), то из характера их прикрепления заметно преимущество последних: плечо действующей силы, развиваемой мышцами при сокращении относительно оси вращения петиолюса, у m. depressor petioli больше, чем у m. elevator petioli superior. Кроме того, опускание может усиливаться действием m. elevator petioli inferior. То же наблюдается и у мышц петиолюса: опускающая мышца явственно длиннее, чем поднимающая, и имеет большое число волокон. Подобное преимущество обусловлено тем, что опускание брюшка, осуществляемое совместным действием m. depressor petioli и m. flexor abdominalis, имеет для![]() Formicinae особое значение. Подгибая брюшко, муравей выбрызгивает яд в ранку, нанесенную челюстями. Примерно так же устроена мускулатура, управляющая движением брюшка у Cataglyphis emeryi и, судя по форме груди и петиолюса, у Polyergus и примитивных представителей Formica (группа
Formicinae особое значение. Подгибая брюшко, муравей выбрызгивает яд в ранку, нанесенную челюстями. Примерно так же устроена мускулатура, управляющая движением брюшка у Cataglyphis emeryi и, судя по форме груди и петиолюса, у Polyergus и примитивных представителей Formica (группа ![]() F. fusca) и Proformica (P. splendida).
F. fusca) и Proformica (P. splendida).
У наиболее продвинутых Formica (подрод Formica s. str) это преимущество опускания еще больше усиливается (рис:24). Основная поверхность проподеума у этих муравьев становится выпуклой, что позволяет расширить площадь прикрепления m. depressor petioli. Верхний край чешуйки приострен и укреплен хитиновым валиком, что повышает жесткость чешуйки в вертикальном направлении. Область прикрепления мышц, опускающих брюшко, распространяется на заднюю поверхность чешуйки, т.е. фактически у каждой мышцы имеются две головки. Кроме того, сами мышцы из-за увеличения высоты чешуйки удлиняются. За счет смещения действующей мышцы к вертикали возрастает угол между вектором силы и направлением на ось. В совокупности все это увеличивает момент действующей силы и соответственно обеспечивает более эффективное подгибание брюшка. Все эти изменения связаны с особенностями защитного поведения Formica s. str. Эти муравьи способны выбрасывать струйку яда (муравьиной кислоты) на большое расстояние, для чего требуется очень сильно подогнуть брюшко.
Совсем иная тенденция наблюдается в пределах рода Cataglyphis. У C. emeryi строение груди и петиолюса и расположение мышц, управляющих движением брюшка, сходно с A. aberrans. У остальных изученных видов рода конечности удлинены. Как уже отмечалось выше, это сопровождается увеличением длины мышц, расположенных в груди, и соответственно изменениями в строении груди. Поскольку мышцы, управляющие движением петиолюса, крепятся к тем же элементам эндоскелета, что и мышцы задних ног, они также удлиняются (рис:20). Это приводит к тому, что увеличивается размах колебаний брюшка. В результате эти муравьи, сохранив способность подгибать брюшко для выбрызгивания яда, приобретают возможность приподнимать его и тем самым несколько смещать центр тяжести. Действительно, все длинноногие Cataglyphis, имеющие нормальную чешуйку (![]() C. aenescens, C. pallida, C. foreli, C. altisquamis), на бегу приподнимают брюшко заметно выше, чем Proformica, Alloformica или C. emeryi. Кроме того, эти муравьи приподнимают брюшко во время отбрасывания почвы передними ногами.
C. aenescens, C. pallida, C. foreli, C. altisquamis), на бегу приподнимают брюшко заметно выше, чем Proformica, Alloformica или C. emeryi. Кроме того, эти муравьи приподнимают брюшко во время отбрасывания почвы передними ногами.
Поскольку как побочный результат изменения формы проподеума возникает новая функция (смещение центра тяжести, повышающее эффективность толчка задними ногами), она может быть подхвачена отбором и усилена. Этот результат может быть получен без изменения уже сложившихся координации между строением конечностей и строением груди за счет перестройки петиолюса и расположенных в нем мышц. В трех группах рода (группы C. bicolor setipes и C. albicans подрода Cataglyphis s. str. и подрод Eumonocombus) независимо возникает узловидный стебелек. Как видно на рис:20, при этом увеличивается поверхность прикрепления m. elevator abdominalis, а сама мышца удлиняется, что обеспечивает большую силу при подъеме брюшка. Одновременно смещается и ось вращения брюшка. В результате эти муравьи приобретают способность держать брюшко вертикально вверх и даже закидывать его на спину (рис:3), максимально перемещая центр тяжести вперед.
Почему такая перестройка не произошла у всех пустынных видов, становится понятно, если рассмотреть строение мышц петиолюса у ![]() Cataglyphis aenescens. На рис:24/2/ видно, что у этого вида, как и у Formica, основание m. depressor abdominalis располагается и на передней, и на задней поверхности чешуйки. По-видимому, у части Cataglyphis (или их предков Alloformica), так и у Formica, начался процесс интенсификации подгибания брюшка. Это, несомненно, было адаптивно, так как позволяло лучше поражать жертву. Однако это закрыло путь, по которому пошли Cataglyphis s. str. и Eumonocombus. Действительно, для того чтобы преобразовать петиолюс типа
Cataglyphis aenescens. На рис:24/2/ видно, что у этого вида, как и у Formica, основание m. depressor abdominalis располагается и на передней, и на задней поверхности чешуйки. По-видимому, у части Cataglyphis (или их предков Alloformica), так и у Formica, начался процесс интенсификации подгибания брюшка. Это, несомненно, было адаптивно, так как позволяло лучше поражать жертву. Однако это закрыло путь, по которому пошли Cataglyphis s. str. и Eumonocombus. Действительно, для того чтобы преобразовать петиолюс типа ![]() C. aenescens в петиолюс типа C. setipes, необходимо вначале вернуться к исходному типу A. aberrans, у которой основание m. depressor abdominalis располагается только на передней поверхности чешуйки (как и у C. setipes). Но такое преобразование ослабит опускание брюшка, не усилив противоположного движения, т.е. приведет к снижению адаптивности.
C. aenescens в петиолюс типа C. setipes, необходимо вначале вернуться к исходному типу A. aberrans, у которой основание m. depressor abdominalis располагается только на передней поверхности чешуйки (как и у C. setipes). Но такое преобразование ослабит опускание брюшка, не усилив противоположного движения, т.е. приведет к снижению адаптивности.
Видимо, такие же перестройки происходили и у южноамериканских, пустынных Dorymyrmex, а также и у древесных![]() Formicinae и
Formicinae и ![]() Dolichoderinae с длинными ногами. Другой возможный путь перемещения центра тяжести и увеличения эффективности толчка — уменьшение относительного размера и веса брюшка. Похоже, что по такому пути пошли южноафриканские Ocymyrmex (рис:11). Но он неизбежно связан со значительной перестройкой топографических координации, так как в брюшке помещаются органы пищеварения и выделения. Такой путь невозможен для
Dolichoderinae с длинными ногами. Другой возможный путь перемещения центра тяжести и увеличения эффективности толчка — уменьшение относительного размера и веса брюшка. Похоже, что по такому пути пошли южноафриканские Ocymyrmex (рис:11). Но он неизбежно связан со значительной перестройкой топографических координации, так как в брюшке помещаются органы пищеварения и выделения. Такой путь невозможен для![]() Formicinae и
Formicinae и ![]() Dolichoderinae, имеющих хорошо развитый зоб, служащий для хранения и транспортировки жидкой пищи.
Dolichoderinae, имеющих хорошо развитый зоб, служащий для хранения и транспортировки жидкой пищи.
Таким образом, ниша, которую занимают в коадаптивном комплексе дневные зоонекрофаги, накладывает характерный отпечаток на облик этих муравьев. Увеличение скорости бега вызывает у них увеличение длины конечностей и изменение пропорций груди. В свою очередь это может повлечь за собой изменение формы стебелька и возникновение способности держать брюшко поднятым вертикально вверх.
Интересно отметить, что обитающие здесь же ночные зоофаги Camponotus в принципе не отличаются морфологически от близких к ним представителей того же рода, обитающих в тропических лесах и гнездящихся в почве. Так, у среднеазиатского C. fedtschenkoi (длина груди (ДГ) равна 2,1—3,4мм) отношение длины заднего бедра к длине груди составляет 0,803±0,014 (n=10), у C. turcestanus (ДГ=3,2—4,2мм) — 0,845±0,009 (n=10) и у C. xerxes (ДГ=3,8—6,0мм) — 0,888±0,014 (n=10). У C. mitis (ДГ=3,0—3,6мм) из Вьетнама этот индекс равен 0,883±0,007 (n=15) и у C. chloroticus из Северной Австралии — 0,849±0,013 (n=10). У близкого к ним C. nicobarensis (ДГ=2,2—3,1мм) из Вьетнама ноги явственно длиннее (индекс равен 0,940±0,016; n=11), так как этот муравей является дендробионтом, хотя и не слишком специализированным. У мелких карпофагов также различий в пропорциях ног не обнаруживается, а у муравьев-жнецов ноги даже относительно короче, чем у крупных Aphaenogaster семиаридных областей.
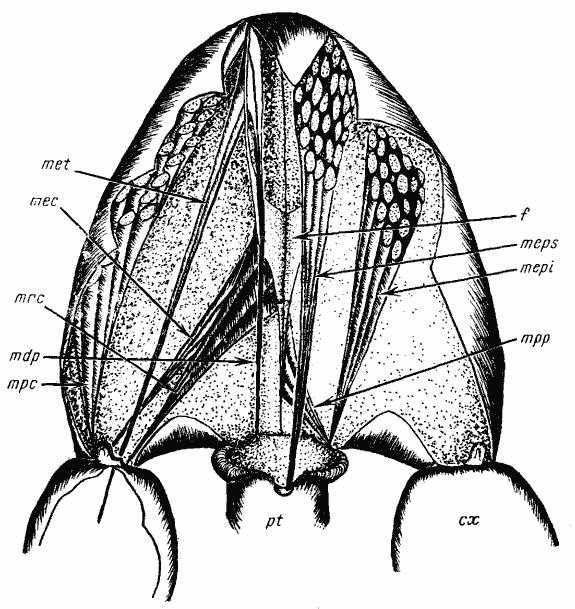
Рис. 18. Мускулатура проподеума Alloformica aberrans сверху
Из каждой пары мышц изображена лишь одна, на левой или правой стороне Обозначения мышц и внутренних скелетных структур (для рис. 18—24): f — фурка проподеума; cx — задняя кокса; mdp — m. depressor petioli; mea — m. elevator abdominalis; mec — m. extensor coxae; mepi — m. elevator petioli interior; meps — m. elevator petioli superior; met — m. extensor trochanters; mfa — m. flexor abdominalis; mpc — m. protractor coxae; mpp — m. protractor petioli; mrc — m. retractor coxae; pt — основание петиолюса
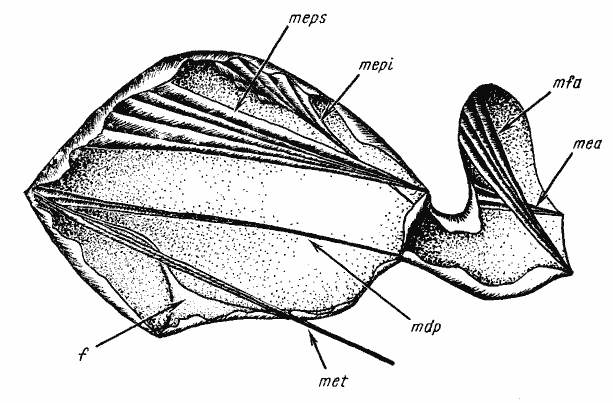
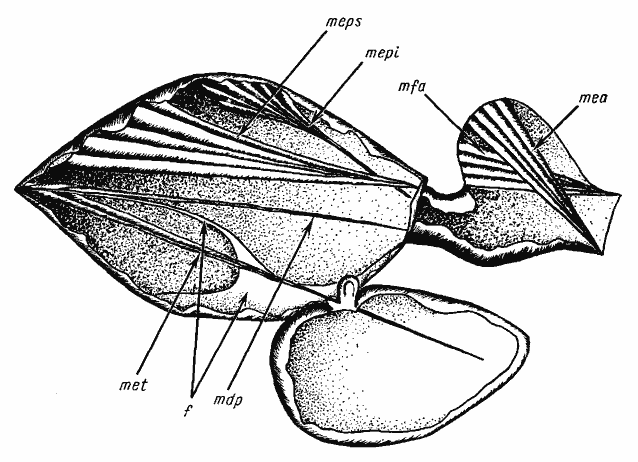

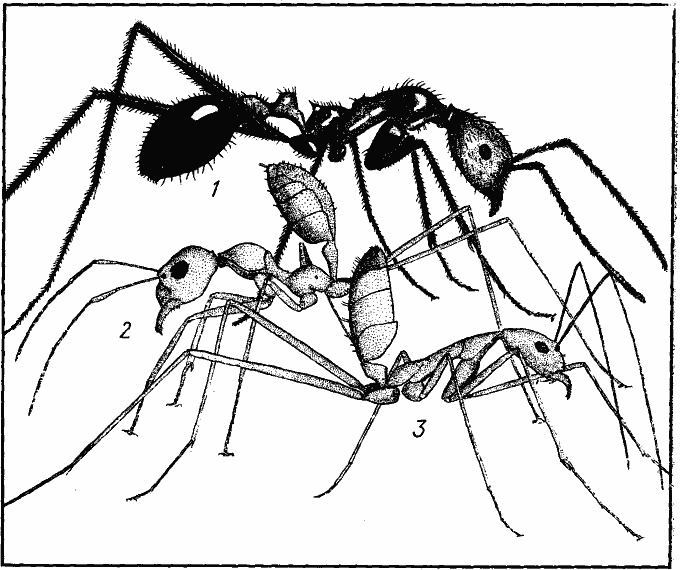
Рис. 22. Некоторые муравьи-дендробионты, добывающие пищу в кронах деревьев
1 — Aphaenogaster swammerdami
2 —
3 — Leptomyrmex fragilis (
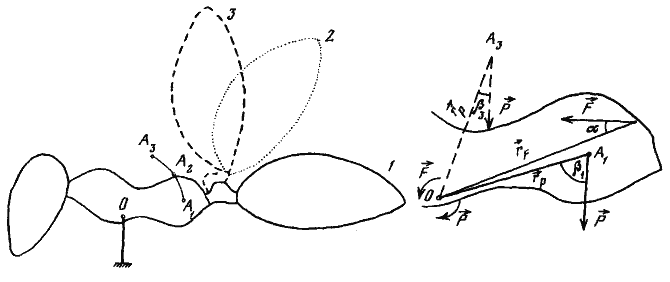
Рис. 23. Схема смещения центра тяжести тела и изменения момента силы тяжести у Cataglyphis setipes при подъеме брюшка
O — ось шарнира опоры; A1 A2, A3 — центр тяжести тела при разных положениях брюшка (1, 2, 3);
P — сила тяжести;
F — сила тяги.
Остальные обозначения в тексте
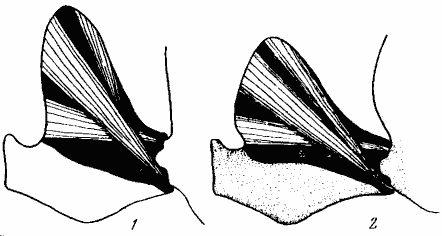
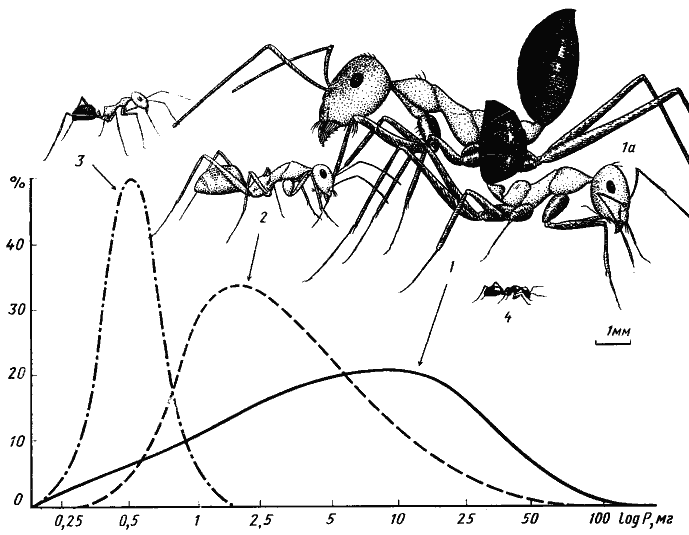
Рис. 3. Сравнительные размеры четырех основных видов дневных зоонекрофагов песчаной пустыни (Акыбай) и частотное распределение (в %) добычи трех видов по весовым классам (усредненные данные)
1, 1а — средний и крупный рабочие Cataglyphis setipes;
2 — C. pallida;
3 — Acantholepis semenovi;
4 — Plagiolepis pallescens
Рис. 11. Дневные зоонекрофаги
1 — Ocymyrmex hirsutus
Всего 65 видов.
Всего 24 вида.
«медовые муравьи»
Всего 22 вида.
Всего 23 вида.
Всего 48 видов.
Всего 17 видов.
Всего 30 видов.
Всего 4 вида.
ксеробионт
Старое название: Proformica deserta
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
ксеробионт
Всего 11 видов.
Всего 100 видов.
Всего 13 видов.
Всего 138 видов.
Включает устаревший: Monacis
Всего 965 видов.
Всего 172 вида.
Включает сокращенный Novomessor.
(лат. mandibule — челюсть, от mando — жую, грызу), первая пара челюстей у ракообразных, многоножек и насекомых. Синоним: жвала.
Myrmicinae — самое большое подсемейство муравьев, включающее более 2 тыс. видов, относящихся более чем к 150 родам.
Всего 32 вида.
Небольшое наиболее архаичное подсемейство муравьев. Распространено по всему земному шару, преимущественно в тропиках. Насчитывает более 230 видов.
Одно из крупных подсемейств, насчитывает около 1,5 тыс. видов, относящихся к 44 родам. Широко распространены на всем земном шаре, довольно обычны в тропиках и занимают доминирующее положение в умеренных широтах.
особый сегмент брюшка. У муравьев бывает двучлениковый (петиолюс + постпетиолюс) и одночлениковый (петиолюс - чешуйка). Количество члеников и форма стебелька очень важны для идентификации вида. Например у муравьев семейства Formicinae стебелек одночлениковый, а у Myrmicinae - двучлениковый.
Всего 17 видов.
организм, обитающий среди растительных или иных органических остатков, на поверхности почвы.
сегмент брюшка.
Всего 6 видов.
 Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant
Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky antполисахарид, образованный остатками аминосахара ацетилглюкозамина. Основной компонент наружного скелета (кутикулы) насекомых, ракообразных и других членистоногих. У грибов заменяет целлюлозу, с которой сходен по химическим и физическим свойствам и биологической роли.
 Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунок
Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунокгигрофил, галобионт
Старое название: Cataglyphis flavigastra
ксеробионт
(зоонекрофаг), организм, питающийся мертвыми животными, гетеротроф
организм, питающийся исключительно другими животными
организм, обитающий в древесном ярусе растительности.
организм, питающийся семенами растений.
Старsые названия: M. hortideorum, M. hortusdeorum
 Oecophylla smaragdina (Fabricius, 1775) — муравей-портной
Oecophylla smaragdina (Fabricius, 1775) — муравей-портной Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравейСтарые названия: Acantholepis semenovi, Acantholepis transcaspiensis
олигогинны
Старое название: Araucomyrmex tener






