






А. Форель (Forel, 1902) и В. М. Вилер (Wheeler, 1908а), а вслед за ними и другие авторы (Goetsch, 1935, 1953; Eisner, Brown, 1958; Wilson, 1971) рассматривают как один из путей адаптации к аридным условиям способность некоторых муравьев сохранять в зобиках особой касты рабочих (плерэргатов или «хранителей меда») большие запасы жидкой пищи. В наибольшей степени эта способность развита у североамериканских Myrmecocystus из группы M. melliger и M. mexicanus и австралийских ![]() Camponotus inflatus (McCook, 1881; Леббок, 1898; Wheeler, 1908а). У плерэргатов этих муравьев склеротизованные тергиты занимают ничтожную часть поверхности раздувшегося брюшка. Эти особи неспособны самостоятельно передвигаться. Более распространены не столь специализированные медовые муравьи, у которых плерэргаты сохраняют способность к самостоятельному движению, хотя и делают это хуже, чем нормальные особи. Склеротизованные тергиты этих плерэргатов занимают приблизительно половину поверхности раздутого брюшка (рис:29). Сюда относятся некоторые южноамериканские Tapinoma, палеарктические Proformica, Prenolepis Палеарктики и Неоарктики, Anoplolepis Южной Африки, Leptomyrmex и Melophorus Австралии (Рузский, 1905; Wheeler, 1908а, 1915а, 1930; Goetsch, 1935; Stumper, 1961; Длусский, 1969а).
Camponotus inflatus (McCook, 1881; Леббок, 1898; Wheeler, 1908а). У плерэргатов этих муравьев склеротизованные тергиты занимают ничтожную часть поверхности раздувшегося брюшка. Эти особи неспособны самостоятельно передвигаться. Более распространены не столь специализированные медовые муравьи, у которых плерэргаты сохраняют способность к самостоятельному движению, хотя и делают это хуже, чем нормальные особи. Склеротизованные тергиты этих плерэргатов занимают приблизительно половину поверхности раздутого брюшка (рис:29). Сюда относятся некоторые южноамериканские Tapinoma, палеарктические Proformica, Prenolepis Палеарктики и Неоарктики, Anoplolepis Южной Африки, Leptomyrmex и Melophorus Австралии (Рузский, 1905; Wheeler, 1908а, 1915а, 1930; Goetsch, 1935; Stumper, 1961; Длусский, 1969а).
Примечательно, что из этого списка два рода (Leptomyrmex и Tapinoma) относятся к подсемейству ![]() Dolichoderinae, а остальные — к подсемейству
Dolichoderinae, а остальные — к подсемейству![]() Formicinae. Медовые муравьи совершенно отсутствуют среди представителей подсемейства
Formicinae. Медовые муравьи совершенно отсутствуют среди представителей подсемейства![]() Myrmicinae, широко представленного во всех пустынях Земли. Предпосылки, обусловившие разное направление эволюции этих муравьев в аридных областях, возникли еще в меловом периоде, при расхождении понероидного и мирмециоидного комплексов подсемейств. С этого момента разными путями пошла эволюция провентрикулюса, запирающего проход из зоба (точнее, из пищевода) в среднюю кишку (Eisner, Brown, 1958). У представителей понероидного комплекса, в частности у
Myrmicinae, широко представленного во всех пустынях Земли. Предпосылки, обусловившие разное направление эволюции этих муравьев в аридных областях, возникли еще в меловом периоде, при расхождении понероидного и мирмециоидного комплексов подсемейств. С этого момента разными путями пошла эволюция провентрикулюса, запирающего проход из зоба (точнее, из пищевода) в среднюю кишку (Eisner, Brown, 1958). У представителей понероидного комплекса, в частности у![]() Myrmicinae, провентрикулюс запирается сфинктором, поэтому для сохранения пиши в зобу необходимо постоянно расходовать энергию. Напротив, у
Myrmicinae, провентрикулюс запирается сфинктором, поэтому для сохранения пиши в зобу необходимо постоянно расходовать энергию. Напротив, у![]() Formicinae и
Formicinae и ![]() Dolichoderinae запирание провентрикулюса осуществляется хитиновыми клапанами, и мышечные усилия нужны для его открывания.
Dolichoderinae запирание провентрикулюса осуществляется хитиновыми клапанами, и мышечные усилия нужны для его открывания.
Неудивительно, что даже в гумидных районах![]() Formicinae и
Formicinae и ![]() Dolichoderinae способны набирать в зоб относительно большее количество жидкости, чем
Dolichoderinae способны набирать в зоб относительно большее количество жидкости, чем![]() Myrmicinae, и соответственно их фуражиры за один рейс могут доставить в гнездо большее количество жидкой пищи. По-видимому, именно эта особенность строения провентрикулюса обусловила высокую степень связи
Myrmicinae, и соответственно их фуражиры за один рейс могут доставить в гнездо большее количество жидкой пищи. По-видимому, именно эта особенность строения провентрикулюса обусловила высокую степень связи![]() Formicinae и
Formicinae и ![]() Dolichoderinae с тлями и другими сосущими насекомыми. Правда, трофобиоз высоко развит и у многих
Dolichoderinae с тлями и другими сосущими насекомыми. Правда, трофобиоз высоко развит и у многих![]() Myrmicinae (например, у Crematogaster), но представители очень многих родов
Myrmicinae (например, у Crematogaster), но представители очень многих родов![]() Myrmicinae (Myrmica, Messor, Leptothorax, Stenamma, Myrmecina и др.) и
Myrmicinae (Myrmica, Messor, Leptothorax, Stenamma, Myrmecina и др.) и![]() Ponerinae (по-видимому, все роды, кроме Odontomachus) либо совсем не посещают тлей, либо делают это эпизодически. У
Ponerinae (по-видимому, все роды, кроме Odontomachus) либо совсем не посещают тлей, либо делают это эпизодически. У![]() Formicinae и
Formicinae и ![]() Dolichoderinae картина обратная: здесь исключение представляют виды, не посещающие сосущих насекомых (пустынные Cataglyphis и Alloformica).
Dolichoderinae картина обратная: здесь исключение представляют виды, не посещающие сосущих насекомых (пустынные Cataglyphis и Alloformica).
Второй предпосылкой возникновения плерэргатов является специфика распределения жидкой пищи в пределах семьи у муравьев гумидных областей. Хорошо известно, что у подавляющего большинства видов муравьев, как и у большинства других общественных насекомых, все члены семьи постоянно обмениваются жидкой пищей. Этот обмен носит название трофаллаксиса. Использование радиоактивных изотопов позволило в 50-х годах показать, что у Formica порция пищи, принесенная в гнездо одним фуражиром, за 20—30 ч равномерно распределяется между 100 и более особями (Gosswald, Kloft, 1956, 1960; Wilson, Eisner, 1957). Впоследствии эти опыты неоднократно повторялись на других видах примерно с такими же результатами.
Для изучения закономерностей распределения пищи в гнезде нами была применена иная методика. Молодая семья Camponotus herculeanus, состоящая из царицы и 45 рабочих, была помещена в гипсовый формикарий. Каждый рабочий имел индивидуальную метку из трех точек, нанесенных на спинку и верх брюшка нитролаками разных цветов.
Во время каждого опыта отмечались все обмены пищей между особями. Направление обмена определялось по различиям в поведении донора и акцептора: во всех случаях донор принимает характерную позу с открытыми жвалами и выдвинутой нижней губой. Опыты длились от 1 до 6 ч. После окончания опытов составлялись детальные схемы пищевых обменов, позволяющие проследить все пути распределения пищи внутри семьи. Наблюдения велись при двух чередующихся режимах:
Режим «А»: в кормушке постоянно находилась пища (мед с водой в пропорции 1:1) и фуражиры постоянно приносили ее в гнездо;
Режим «В»: пища в кормушке отсутствовала и в гнезде распределялась пища, запасенная в зобиках рабочих.
При режиме «А» в среднем за 15 мин в гнезде происходит 19,2±2,1 обменов пищей, а при режиме «В» — в 1,5 раза меньше (12,6±1,6 обменов/15 мин). Когда семья получает пищу после длительного голодания, интенсивность обменов вначале резко возрастает, а затем медленно падает ниже нормы (рис:30/1/). При постоянном режиме «А» наблюдаются постоянные колебания интенсивности обмена с периодом колебаний 1,5—2 ч.
На рис:30/2/ в качестве примера показано распределение в семье трех порций пищи, принесенных тремя фуражирами. Характер кривых мало отличается от характера суммарной кривой, показывающей рост числа рабочих, получивших пищу из кормушки: скорость распределения пищи, высокая вначале, постепенно падает. Это происходит в результате того, что при высокой интенсивности обмена и ограниченности группы активных рабочих (всего 19 особей из 45) быстро нарастает число повторных обменов между особями.
При режиме «В» распределение носит иной характер. Пища из зобика одного рабочего вначале распространяется в семье медленно, но постоянно происходит постепенное нарастание скорости (рис:30/3/), поскольку размер обменивающейся группы увеличивается.
Если наблюдать за поведением муравьев при трофаллаксисе в течение нескольких часов подряд, можно заметить, что функции одних и тех же рабочих при двух разных пищевых режимах различны. 13 особей (29%) практически не участвовали в трофаллаксисе. Из особей, которые были активны хотя бы при одном пищевом режиме, подавляющее большинство (29 из 32) сменяли функции при смене режимов. 13 из 23 особей, активных при режиме «В», не были активны при режиме «А».
Особенно резко видны различия при сравнении функциональных доноров. При режиме «В» ими становились особи, которые при режиме «А» либо не были активными, либо были акцепторами. Таким образом в зобиках этих муравьев происходило накопление пищи при режиме «А», а при режиме «В» они являлись основными хранителями пищи в семье и распределяли ее между другими ее членами.
На основании этих опытов можно утверждать, что у муравьев гумидных областей может одновременно существовать два пути распределения пищи: распределение пищи, приносимой фуражирами, и распределение пищи, сохраняемой в зобиках рабочих-«накопителей». Второй процесс особенно активно происходит через несколько дней после прекращения поступления пищи извне. Характер и скорость распределения при этих двух процессах различны, а одни и те же рабочие выполняют различные, часто противоположные функции.
Хотя количество пищи, запасаемой плерэргатами, значительно больше, чем у рабочих-«накопителей», общий характер их поведения в семье практически не отличается. Как показал Р. Штумпер (Stumper, 1961), у европейского медового муравья Proformica nasuta плерэргаты при обычных условиях являются акцепторами и основное направление движения пищи происходит от фуражиров к плерэргатам. При повышении температуры до 30—31°, когда резко возрастают энергетические траты семьи, или при прекращении поступления пищи извне плерэргаты становятся донорами и ток пищи в семье меняется на обратный.
Значение плерэргатов для жизни, семьи очевидно. В аридных и субаридных областях всегда бывают довольно длительные периоды, когда муравьи практически не могут добыть пищи. Мед, хранящийся в плерэргатах Myrmecocystus, содержит от 99 до 45% воды. От 57 до 63% сухого остатка составляют углеводы (глюкоза, фруктоза, мальтоза, сахароза), остальное приходится главным образом на липиды и глицерин. Аминокислоты составляют лишь очень небольшую долю сухого остатка (около 3%), так что «мед» не может быть использован для выращивания расплода и может обеспечивать только восполнение энергетических и водных затрат взрослых муравьев (Burgett, Young, 1974; Conway, 1977). У Myrmecocystus mexicanus брюшко плерэргата содержит около 0,39 г «меда», что составляет 89% веса тела (McCook, 1881). Поскольку крупные семьи Myrmecocystus содержат 500—1500 плерэргатов (McCook, 1881; Creighton, Crandall, 1954; Conway, 1977), их запасы должны составлять 200—600 г «меда».
Г. Мак-Кук содержал группы из рабочих и плерэргатов M. mexicanus в течение четырех месяцев без всякой пищи. Как он пишет, «после опыта рабочие имели хорошо наполненное брюшко, как фуражиры, возвращающиеся с добычей. Хранители меда несколько похудели, но все же отличались размерами брюшка от рабочих» (McCook, 1881). У Proformica nasuta содержимое зобика одного плерэргата (10 мг) может обеспечить питанием 100 рабочих в течение 30 дней (Stumper, 1961).
В пустынях Средней Азии и Северной Африки настоящие медовые муравьи отсутствуют. Лишь в северные пустыни Палеарктики (Казахстан, Монголия) заходят некоторые степные виды Proformica. В Средней Азии лишь три вида муравьев (Cataglyphis pallida, C. cinnamomea, Camponotus xerxes) способны запасать достаточно большое количество жидкой пищи в зобиках крупных рабочих. Плерэргаты этих видов, если их можно так назвать, явственно отличаются от нормальных рабочих, но у них при заполненном брюшке межсегментные участки растянутой кутикулы уже, чем тергиты. Так же выглядят плерэргаты Camponotus thoracicus из Центральной Сахары (Delye, 1968) и, судя по рисунку, Melophorus bagoti из пустынь Австралии (Wheeler, 1908а). У C. pallida средний вес рабочих в верхних камерах гнезда (главным образом это фуражиры) при весенней раскопке в Акыбае в 1971 г. составлял 1,37 мг, а плерэргатов (при тех же размерах) — 1,94 мг, т.е. рабочие этого вида могут сохранять в зобу 0,6—0,7 мг жидкой пищи, что составляет 31—36% от веса тела. В двух раскопанных гнездах (№71—08 и 71—10) общее число рабочих было 1193 и 756, в том числе плерэргатов соответственно 170 (14,2%) и 35 (4,6%). В целом запасы жидкой пищи в гнезде №71—08 были 100—120 мг и в гнезде №71—10 — 20—25 мг. Около 30% веса тела содержимое зоба составляет также у Camponotus thoracicus (Delye, 1968), C. xerxes и Cataglyphis cinnamomea. У остальных муравьев пустынь Средней Азии и Сахары содержимое заполненного зоба, как и у муравьев гумидных областей, не превышает 10—20% от веса тела.
Несомненно, способность муравьев делать запасы жидкой пищи дает им в пустынях явное преимущество. В апреле 1971 г. в Акыбае мы поставили следующий опыт: в чашки Петри, застеленные фильтровальной бумагой, помещалось по 20 рабочих Cataglyphis pallida, ![]() C. aenescens flavigastra и Messor aralocaspius. У первого вида вес наполненного зоба составляет более 30% веса тела, у второго — около 20% и у третьего — около 15%. Муравьи в течение суток перед опытом выдерживались в камере, где находилась кормушка с сахарным сиропом, и зобики всех особей были заполнены. В течение опыта муравьи получали только воду. Каждый опыт был сделан в трех повторностях. Через 20 суток после начала опыта погибли все рабочие M. aralocaspius и 88% рабочих
C. aenescens flavigastra и Messor aralocaspius. У первого вида вес наполненного зоба составляет более 30% веса тела, у второго — около 20% и у третьего — около 15%. Муравьи в течение суток перед опытом выдерживались в камере, где находилась кормушка с сахарным сиропом, и зобики всех особей были заполнены. В течение опыта муравьи получали только воду. Каждый опыт был сделан в трех повторностях. Через 20 суток после начала опыта погибли все рабочие M. aralocaspius и 88% рабочих ![]() C. aenescens. В то же время через 30 суток в живых осталось больше половины (57%) рабочих C. pallida (рис:31). Различия в гибели нельзя объяснить различной теплоустойчивостью муравьев, так как хотя во время опыта температура менялась, она никогда не превышала 26°. Расход пищи в первую неделю опыта также был приблизительно одинаков: у C. pallida он составлял 0,0642 мг/сут, или 2,25% от исходного веса тела, а у M. aralocaspius — 0,158 мг/сут, или 2,9% от исходного веса тела. Средний вес рабочих C. pallida в начале опыта равнялся 2,87 мг, через неделю он упал до 2,42 мг, через две недели — до 2,30 мг и через месяц — до 2,19 мг. В отличие от солдат плерэргаты не являются морфологической кастой. Их анатомическое строение не отличается от строения нормальных рабочих такого же размера (сравнивается, естественно, размер жестких структур, например головной капсулы), выполняющих обычные работы в гнезде (McCook, 1881; Леббок, 1898). По-видимому, плерэргатом потенциально может стать любой крупный молодой рабочий. Как это ни удивительно, но выработка способности к сохранению в зобу больших количеств жидкости вызывает гораздо меньшее изменение сложившихся координации, чем описанное выше увеличение скорости бега. По сути дела, необходимо лишь повысить растяжимость межсегментной кутикулы брюшка и стенок зоба. Единственное заметное морфологическое изменение, иногда наблюдающееся у медовых муравьев, — искривление задних бедер (рис:32). Строение провентикулюса медовых муравьев в принципе не отличается от строения у близких родов, не имеющих плерэргатов (Eisner, 1957). Возникает вопрос, почему же при явных преимуществах в аридном климате и при наличии морфологических предпосылок настоящие медовые муравьи не возникли в пустынях Палеарктики? В Неоарктике также ночные Myrmecocystus с настоящими плерэргатами приурочены больше к семиаридным областям, а в пустынях встречаются в основном там, где имеется сравнительно обильная древесно-кустарниковая растительность (Wheeler, 1908а). Кажется, единственной причиной этого является отсутствие обильных источников жидкой углеводной пищи. Для того чтобы пояснить эту мысль, приведем некоторые ориентировочные подсчеты, сделанные для Camponotus xerxes. В пустынях мне никогда не встречалось большего количества тлей, выделяющих падь, чем в саксауловых лесах Акыбая на границе с оазисом. Но и в этих условиях рабочие-трофобионты гнезда №71—31, собиравшие падь тлей Brachyunguis saxaulica и червецов Pulvinaria orientalis на саксаулах, расположенных в нескольких метрах от гнезда, тратили на сбор пади в среднем около 5 ч и за ночь совершали в среднем 2,05 рейса. Для сравнения можно отметить, что в лесной зоне рабочие-трофобионты
C. aenescens. В то же время через 30 суток в живых осталось больше половины (57%) рабочих C. pallida (рис:31). Различия в гибели нельзя объяснить различной теплоустойчивостью муравьев, так как хотя во время опыта температура менялась, она никогда не превышала 26°. Расход пищи в первую неделю опыта также был приблизительно одинаков: у C. pallida он составлял 0,0642 мг/сут, или 2,25% от исходного веса тела, а у M. aralocaspius — 0,158 мг/сут, или 2,9% от исходного веса тела. Средний вес рабочих C. pallida в начале опыта равнялся 2,87 мг, через неделю он упал до 2,42 мг, через две недели — до 2,30 мг и через месяц — до 2,19 мг. В отличие от солдат плерэргаты не являются морфологической кастой. Их анатомическое строение не отличается от строения нормальных рабочих такого же размера (сравнивается, естественно, размер жестких структур, например головной капсулы), выполняющих обычные работы в гнезде (McCook, 1881; Леббок, 1898). По-видимому, плерэргатом потенциально может стать любой крупный молодой рабочий. Как это ни удивительно, но выработка способности к сохранению в зобу больших количеств жидкости вызывает гораздо меньшее изменение сложившихся координации, чем описанное выше увеличение скорости бега. По сути дела, необходимо лишь повысить растяжимость межсегментной кутикулы брюшка и стенок зоба. Единственное заметное морфологическое изменение, иногда наблюдающееся у медовых муравьев, — искривление задних бедер (рис:32). Строение провентикулюса медовых муравьев в принципе не отличается от строения у близких родов, не имеющих плерэргатов (Eisner, 1957). Возникает вопрос, почему же при явных преимуществах в аридном климате и при наличии морфологических предпосылок настоящие медовые муравьи не возникли в пустынях Палеарктики? В Неоарктике также ночные Myrmecocystus с настоящими плерэргатами приурочены больше к семиаридным областям, а в пустынях встречаются в основном там, где имеется сравнительно обильная древесно-кустарниковая растительность (Wheeler, 1908а). Кажется, единственной причиной этого является отсутствие обильных источников жидкой углеводной пищи. Для того чтобы пояснить эту мысль, приведем некоторые ориентировочные подсчеты, сделанные для Camponotus xerxes. В пустынях мне никогда не встречалось большего количества тлей, выделяющих падь, чем в саксауловых лесах Акыбая на границе с оазисом. Но и в этих условиях рабочие-трофобионты гнезда №71—31, собиравшие падь тлей Brachyunguis saxaulica и червецов Pulvinaria orientalis на саксаулах, расположенных в нескольких метрах от гнезда, тратили на сбор пади в среднем около 5 ч и за ночь совершали в среднем 2,05 рейса. Для сравнения можно отметить, что в лесной зоне рабочие-трофобионты ![]() Formica rufa возвращаются в гнездо с заполненным зобом в среднем через 2 ч. Лишь у 29% фуражиров (74 из 254) C. xerxes, возвращающихся в гнездо, брюшко было заметно раздуто. По ориентировочным подсчетам /Разница между средним весом фуражиров, возвращающихся в гнездо с раздувшимся брюшком и выходящих из гнезда, составляет около 5,5 мг; для остальных фуражиров, возвращающихся в гнездо без груза, мы приняли, что вес содержимого их зоба составляет 10% от веса тела, или 2,3 мг, так как при таком заполнении брюшка еще трудно отличить муравьев с заполненным зобом./, фуражиры этого гнезда приносили в один вход ежедневно не более 800 мг пади. Поскольку тли в Каракумах активны практически два месяца (апрель — май), за год это составит около 48 г. Данные раскопок других гнезд показали, что у C. xerxes на один вход приходится около 600 рабочих общим весом 19 г. Приняв на основании описанного выше опыта с голоданием, что каждый рабочий расходует (в спокойном состоянии) содержимое зоба, равное 3% веса его тела, мы получим, что за те же два месяца муравьи израсходуют 28 г пади. Остаток составит всего 20 г, или менее чем двухмесячный запас. На самом деле это количество будет еще меньше, так как, во-первых, активно двигающиеся фуражиры будут расходовать больше энергии, во-вторых, концентрация сахаров в содержимом зоба у муравьев выше, чем в пади, и, в-третьих, часть сахара будет перерабатываться и откладываться в виде жира (у рабочих пустынных Camponotus жировое тело обычно хорошо развито).
Formica rufa возвращаются в гнездо с заполненным зобом в среднем через 2 ч. Лишь у 29% фуражиров (74 из 254) C. xerxes, возвращающихся в гнездо, брюшко было заметно раздуто. По ориентировочным подсчетам /Разница между средним весом фуражиров, возвращающихся в гнездо с раздувшимся брюшком и выходящих из гнезда, составляет около 5,5 мг; для остальных фуражиров, возвращающихся в гнездо без груза, мы приняли, что вес содержимого их зоба составляет 10% от веса тела, или 2,3 мг, так как при таком заполнении брюшка еще трудно отличить муравьев с заполненным зобом./, фуражиры этого гнезда приносили в один вход ежедневно не более 800 мг пади. Поскольку тли в Каракумах активны практически два месяца (апрель — май), за год это составит около 48 г. Данные раскопок других гнезд показали, что у C. xerxes на один вход приходится около 600 рабочих общим весом 19 г. Приняв на основании описанного выше опыта с голоданием, что каждый рабочий расходует (в спокойном состоянии) содержимое зоба, равное 3% веса его тела, мы получим, что за те же два месяца муравьи израсходуют 28 г пади. Остаток составит всего 20 г, или менее чем двухмесячный запас. На самом деле это количество будет еще меньше, так как, во-первых, активно двигающиеся фуражиры будут расходовать больше энергии, во-вторых, концентрация сахаров в содержимом зоба у муравьев выше, чем в пади, и, в-третьих, часть сахара будет перерабатываться и откладываться в виде жира (у рабочих пустынных Camponotus жировое тело обычно хорошо развито).
Отсюда следует, что даже при благоприятных условиях содержимое зоба, накопленное за время активности сосущих насекомых, не может служить основным источником восполнения энергетических трат C. xerxes в летние и осенние месяцы, когда рабочие этого вида продолжают фуражировку и строительную деятельность. В таких условиях наличие развитых плерэргатов не может давать существенных преимуществ и вполне достаточной оказывается способность накапливать в зобу запасы, составляющие не более 30—40% от веса тела. Пустынные Cataglyphis вообще утратили связь с тлями. На то, что это скорее всего именно вторичная утрата, указывает, во-первых, то, что строение их провентикулюса ничуть не примитивнее, чем у Formica, а во-вторых, то, что европейские ![]() C. aenescens посещают тлей так же, как Serviformica.
C. aenescens посещают тлей так же, как Serviformica.
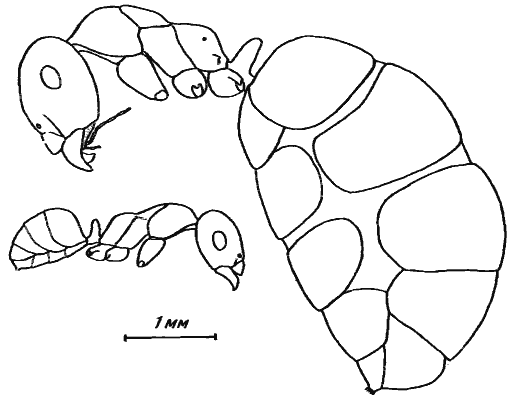
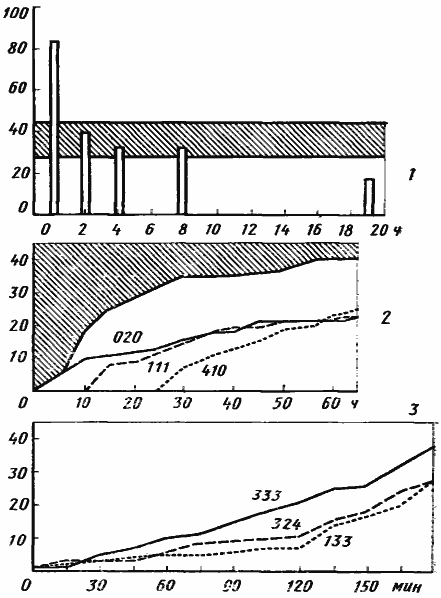
Рис. 30. Распределение жидкой пищи в лабораторной семье Camponotus herculeanus, состоящей из самки, 45 рабочих и расплода (100—150 личинок I возраста)
1 — изменение интенсивности трофаллаксисов (число трофаллаксисов за 30 мин) при переходе к режиму «А» после длительного голодания. 27—28 сентября 1966 г. Заштрихованная полоса — пределы, в которых колебалась интенсивность трофаллаксисов в течение 2 ч до начала опыта;
2 — распределение жидкой пищи внутри семьи при пищевом режиме «А» (тот же опыт, что на верхнем рисунке). Кривые показывают нарастание числа рабочих, получивших часть одной порции пищи, принесенной фуражирами №№020, 111 и 410. Заштриховано — число рабочих, не получивших пищи из кормушки в течение первого часа опыта;
3 — распределение жидкой пищи при пищевом режиме «В». 11 сентября 1966 г. В течение трех дней семья не получала пищи извне. Кривые показывают нарастание числа рабочих, получивших пищу из зобиков функциональных доноров №№ 133, 324 и 333
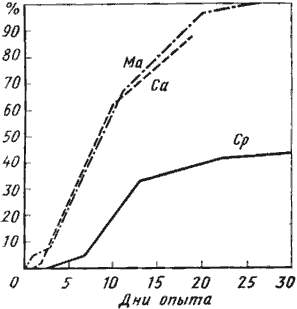
Рис. 31. Динамика гибели муравьев (в %) при длительном голодании
Перед началом опыта зобики всех муравьев были заполнены сахарным сиропом.
Ca —
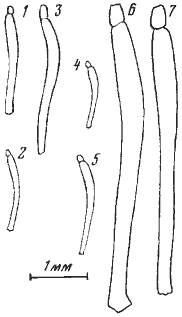
Рис. 32. Бедра задних ног (сверху) некоторых представителей трибы Formicini
1 — Alloformica aberrans; 2 — Proformica epinotalis (крупный рабочий); 3 —
у муравьев имеется три основные касты: самцы, самки и рабочие (бесплодные модифицированные самки).
рабочие с сильно раздутым брюшком; это каста, так называемых, "медовых бочек".
Всего 30 видов.
Старsые названия: M. hortideorum, M. hortusdeorum
 Camponotus inflatus Lubbock, 1880 — медовый муравей
Camponotus inflatus Lubbock, 1880 — медовый муравейсегмент брюшка.
Всего 62 вида.
Всего 24 вида.
Всего 11 видов.
«медовые муравьи»
Всего 22 вида.
Всего 17 видов.
Всего 23 вида.
Небольшое наиболее архаичное подсемейство муравьев. Распространено по всему земному шару, преимущественно в тропиках. Насчитывает более 230 видов.
Одно из крупных подсемейств, насчитывает около 1,5 тыс. видов, относящихся к 44 родам. Широко распространены на всем земном шаре, довольно обычны в тропиках и занимают доминирующее положение в умеренных широтах.
Myrmicinae — самое большое подсемейство муравьев, включающее более 2 тыс. видов, относящихся более чем к 150 родам.
необратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.
полисахарид, образованный остатками аминосахара ацетилглюкозамина. Основной компонент наружного скелета (кутикулы) насекомых, ракообразных и других членистоногих. У грибов заменяет целлюлозу, с которой сходен по химическим и физическим свойствам и биологической роли.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
взаимовыгодные отношения между муравьями и выделяющими падь насекомыми: слизывая падь, муравьи охраняют этих насекомых.
Всего 427 видов.
Всего 106 видов.
Всего 19 видов.
Всего 50 видов.
Всего 26 видов.
Довольно примитивная группа муравьев по своему строению и поведению. Рабочие мономорфны, самки по размерам близки к рабочим.
Всего 65 видов.
Всего 4 вида.
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
живут колониями (пчелы, муравьи, термиты, некоторые осы). В состав колонии, напр. у термитов, входят половые особи (самцы и матки обеспечивают размножение), солдаты (защиту) и рабочие (собирание корма, строительство гнезда и т. д.). Солдаты и рабочие бесплодны; половые особи и солдаты сами не способны питаться; таким образом, члены колонии полностью зависят друг от друга. Разделение функций и сложные взаимодействия разных особей в колониях общественных насекомых обеспечиваются рядом особенностей в их строении и поведении; последнее регулируется с помощью выделяемых ими химических веществ (феромонов).
(от трофо… и греч. allaxis — обмен) передача пищи и гормональных веществ от особи к особи путем кормления содержимым зобика, желудка или слизывания выпота с поверхности тела. Играет огромную роль в передаче информации между муравьями.
Всего 965 видов.
искусственный муравейник.
(лат. acceptor - приемник) - принимающий на себя.
(лат. mandibule — челюсть, от mando — жую, грызу), первая пара челюстей у ракообразных, многоножек и насекомых. Синоним: жвала.
Старое название: Cataglyphis cinnamomea
 Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунок
Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунокгигрофил, галобионт
Старое название: Cataglyphis flavigastra
ксеробионт
сладкая густая жидкость, выделяемая тлями, червецами и другими насекомыми, питающимися соком растений. Медоносная пчела собирает падь с листьев и перерабатывает в падевый мед.
общее название нескольких семейств насекомых подотряда кокцидовых. Св. 1600 видов, преимущественно в тропиках и субтропиках. У тропических лаковых червецов покровы образованы застывающими на воздухе лаковыми выделениями. Многие (напр., мучнистые червецы) вредители растений, особенно субтропических (напр., цитрусовых).
 Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравейУстарел, исключен.
ксеробионт
Старое название: Proformica deserta
ксеробионт
ксеробионт






