






В. М. Вилер (Wheeler, 1907а) обратил внимание на то, что у многих пустынных муравьев параллельно возникают характерные длинные щетинки, названные им аммохетами, никогда не встречающиеся у видов гумидных областей. Часто они располагаются правильными рядами. Аммохеты наличника, жвал и нижней стороны головы всегда образуют вместе некоторое подобие корзинки. Однако Вилер не смог понять значения этих органов, так как вел наблюдения в искусственных гипсовых гнездах, где муравьи не рыли почвы. Он предположил, что они служат для переноски воды. Вскоре Ф. Санчи (Santschi, 1909) обнаружил, что в естественных условиях муравьи используют «корзинку», образованную аммохетами и нижнечелюстными щупиками, для переноски почвы. Этот орган он назвал псаммофором. Почти сразу же эти наблюдения были подтверждены А. Форелем (Forel, 1911) и В. М. Вилером (Wheeler., 1912).
В настоящее время псаммофоры известны и описаны у представителей родов Messor, Pogonomyrmex, Veromessor, Novomessor, Monomorium, Chelaner, Goniomma, Oxyopomyrmex, Ocymyrmex, Conomyrma, Dorymyrmex, Araucomyrmex, Camponotus, Melophorus, Cataglyphis, Myrmecocystus (Wheeler, 1907a; Santschi, 1909; Emery, 1910—1925; Spangler, Rettenmeyer, 1966; Snelling, 1973; Арнольди, 1977). В состав псаммофора у разных видов муравьев могут входить нижнечелюстные щупики, щетинки на наличнике (клипеальные аммохеты), на жвалах (мандибулярные аммохеты), на ментуме нижней губы (ментальные аммохеты) и на нижней поверхности головы (гулярные аммохеты). Нижнечелюстные щупики, если они участвуют в переноске комка почвы, имеют удлиненные, а иногда вдобавок уплощенные 3-й и 4-й членики и обычно покрыты длинными волосками.
Высоко специализированные псаммофоры разных представителей подсемейства![]() Myrmicinae (Ocymyrmex, наиболее специализированные Messor, Pogonomyrmex и Monomorium) устроены довольно сходно. Дно и заднюю стенку его «корзины» образует ряд длинных гулярных аммохет, расположенных полукругом. Площадка на нижней стороне головы, которую они окружают, часто гладкая и блестящая, тогда как остальная поверхность головы может быть скульптированной. Боковые стенки псаммофора образуют жвалы с мандибулярными аммохетами, а переднюю — аммохеты, расположенные на переднем крае наличника (рис:13/1,2,4,5/; рис:15). В псаммофорах
Myrmicinae (Ocymyrmex, наиболее специализированные Messor, Pogonomyrmex и Monomorium) устроены довольно сходно. Дно и заднюю стенку его «корзины» образует ряд длинных гулярных аммохет, расположенных полукругом. Площадка на нижней стороне головы, которую они окружают, часто гладкая и блестящая, тогда как остальная поверхность головы может быть скульптированной. Боковые стенки псаммофора образуют жвалы с мандибулярными аммохетами, а переднюю — аммохеты, расположенные на переднем крае наличника (рис:13/1,2,4,5/; рис:15). В псаммофорах![]() Myrmicinae примитивного типа (Goniomma, Oxyopomyrmex, Novomessor, многие Messor, Monomorium и Pogonomyrmex) гулярные аммохеты образуют «бороду», но не располагаются правильным рядом, а равномерно распределены по всей нижней поверхности головы (рис:13/3/). Основная нагрузка при переноске песка приходится на гулярные и мандибулярные аммохеты, тогда как клипеальные лишь придерживают песчинки. Как показали эксперименты с Pogonomyrmex (Spangler, Rettenmeyer, 1966), при удалении аммохет муравьи продолжают стереотипное поведение. Но при удалении гулярных аммохет производительность переноски мелкого (размер частиц 0,5—0,9 мм) песка снижается в 3—4 раза, а при удалении клипеальных практически не меняется.
Myrmicinae примитивного типа (Goniomma, Oxyopomyrmex, Novomessor, многие Messor, Monomorium и Pogonomyrmex) гулярные аммохеты образуют «бороду», но не располагаются правильным рядом, а равномерно распределены по всей нижней поверхности головы (рис:13/3/). Основная нагрузка при переноске песка приходится на гулярные и мандибулярные аммохеты, тогда как клипеальные лишь придерживают песчинки. Как показали эксперименты с Pogonomyrmex (Spangler, Rettenmeyer, 1966), при удалении аммохет муравьи продолжают стереотипное поведение. Но при удалении гулярных аммохет производительность переноски мелкого (размер частиц 0,5—0,9 мм) песка снижается в 3—4 раза, а при удалении клипеальных практически не меняется.
Гораздо разнообразнее строение псаммофоров у представителей подсемейств![]() Formicinae и
Formicinae и ![]() Dolichoderinae. У Camponotus, как и у
Dolichoderinae. У Camponotus, как и у![]() Myrmicinae, его образуют гулярные, клипеальные и мандибулярные аммохеты (рис:16/1/). Как показали наши наблюдения за поведением C. turcestanicus, нижнечелюстные щупики этого муравья не принимают участия в переноске комка почвы и во время работы плотно прижаты к голове. Среди представителей рода наиболее совершенный псаммофор описан у южноафриканского C. mystaceus. У этого вида клипеальные аммохеты располагаются не на переднем крае, как у всех остальных муравьев, а образуют правильный ряд в задней половине наличника (рис:16/2/). У североамериканских Myrmecocystus заднюю стенку и дно «корзины» образуют гулярные аммохеты, переднюю — клипеальные аммохеты, а бока (и частично дно) — мандибулы и удлиненные нижнечелюстные щупики, покрытые длинными щетинками (рис:16/7/). Аналогичное строение имеет псаммофор южноамериканских Dorymyrmex (подсемейство
Myrmicinae, его образуют гулярные, клипеальные и мандибулярные аммохеты (рис:16/1/). Как показали наши наблюдения за поведением C. turcestanicus, нижнечелюстные щупики этого муравья не принимают участия в переноске комка почвы и во время работы плотно прижаты к голове. Среди представителей рода наиболее совершенный псаммофор описан у южноафриканского C. mystaceus. У этого вида клипеальные аммохеты располагаются не на переднем крае, как у всех остальных муравьев, а образуют правильный ряд в задней половине наличника (рис:16/2/). У североамериканских Myrmecocystus заднюю стенку и дно «корзины» образуют гулярные аммохеты, переднюю — клипеальные аммохеты, а бока (и частично дно) — мандибулы и удлиненные нижнечелюстные щупики, покрытые длинными щетинками (рис:16/7/). Аналогичное строение имеет псаммофор южноамериканских Dorymyrmex (подсемейство ![]() Dolichoderinae). Однако у этих муравьев щетинки на нижнечелюстных щупиках невелики, но 3-й и 4-й членики щупиков резко удлинены и уплощены, а 5-й членик причленяется не к вершине, а к внешней поверхности 4-го (рис:16/10,11/). По-видимому, псаммофор Dorymyrmex является самым емким и совершенным среди муравьев вообще. У представителей другого пустынного рода
Dolichoderinae). Однако у этих муравьев щетинки на нижнечелюстных щупиках невелики, но 3-й и 4-й членики щупиков резко удлинены и уплощены, а 5-й членик причленяется не к вершине, а к внешней поверхности 4-го (рис:16/10,11/). По-видимому, псаммофор Dorymyrmex является самым емким и совершенным среди муравьев вообще. У представителей другого пустынного рода ![]() Dolichoderinae — североамериканских Conomyrma псаммофор развит слабо, но все же, как это показал Снеллинг (Snelling, 1974), имеется.
Dolichoderinae — североамериканских Conomyrma псаммофор развит слабо, но все же, как это показал Снеллинг (Snelling, 1974), имеется.
Еще один тип псаммофора наблюдается у палеарктических Cataglyphis и австралийских Melophorus. У этих муравьев дно и заднюю стенку «корзины» образует пучок расходящихся ментальных аммохет и удлиненные нижнечелюстные щупики, покрытые длинными аммохетами, направленными вперед и назад. Боковые и передняя стенки, как и у других муравьев, образованы клипеальными и мандибулярными аммохетами (рис:16/5,6/).
Широко распространено мнение, что псаммофор появляется у пустынных муравьев в связи с необходимостью переносить сухую почву (Santschi, 1909; Spangler, Rettenmeyer, 1966; Delye, 1968). Это подтверждается и нашими лабораторными экспериментами. Описанные выше опыты Спанглера и Реттенмейера страдают серьезным методическим недостатком: муравьи с ампутированными аммохетами могли вести себя иначе, чем нормальные особи, хотя исследователи этого не заметили. Поэтому мы использовали другой прием и попытались сравнить эффективность выноса почвы у представителей близких родов, один из которых имеет развитый псаммофор (![]() Cataglyphis aenescens), а у другого псаммофор отсутствует (Alloformica aberrans). В узкостенный садок с увлажненной песчаной почвой было вначале помещено 10 рабочих A. aberrans. Длина груди этих особей была 1,5—2,1 мм (среднее—1,85 мм), а вес равен 2,3—5,8 мг (среднее — 4,0 мг). Муравьи выносили на поверхность слепленные лапками и головой комочки почвы в челюстях. Нижнечелюстные щупики при этом комка не поддерживали и либо были прижаты к нижней стороне головы, либо свободно висели. Средний вес выносившихся комков почвы был равен 0,94 мг. На следующий день в тот же садок было помещено 10 рабочих
Cataglyphis aenescens), а у другого псаммофор отсутствует (Alloformica aberrans). В узкостенный садок с увлажненной песчаной почвой было вначале помещено 10 рабочих A. aberrans. Длина груди этих особей была 1,5—2,1 мм (среднее—1,85 мм), а вес равен 2,3—5,8 мг (среднее — 4,0 мг). Муравьи выносили на поверхность слепленные лапками и головой комочки почвы в челюстях. Нижнечелюстные щупики при этом комка не поддерживали и либо были прижаты к нижней стороне головы, либо свободно висели. Средний вес выносившихся комков почвы был равен 0,94 мг. На следующий день в тот же садок было помещено 10 рабочих ![]() C. aenescens. Специально были отобраны особи, линейные размеры которых были такими же, как у A. aberrans (длина груди равна 1,6—2,2 мм, среднее—1,83 мм). Однако, поскольку
C. aenescens. Специально были отобраны особи, линейные размеры которых были такими же, как у A. aberrans (длина груди равна 1,6—2,2 мм, среднее—1,83 мм). Однако, поскольку ![]() C. aenescens имеют более стройное тело, вес муравьев был вдвое меньше: 0,8—3,6 мг, среднее — 2,3 мг. Хотя эти муравьи и пользовались псаммофором, средний вес выносимых комков почвы практически не отличался и составлял 0,98 мг. Затем муравьи были последовательно помещены в садок с сухой почвой.
C. aenescens имеют более стройное тело, вес муравьев был вдвое меньше: 0,8—3,6 мг, среднее — 2,3 мг. Хотя эти муравьи и пользовались псаммофором, средний вес выносимых комков почвы практически не отличался и составлял 0,98 мг. Затем муравьи были последовательно помещены в садок с сухой почвой. ![]() C. aenescens продолжали рыть, и эффективность их рытья заметно не изменилась (взвешивание не производилось, но об этом можно было приблизительно судить по размерам комков). Помещенные в тот же садок A. aberrans вначале пытались отгребать почву лапками и головой, но не переносили ее в челюстях, а вскоре прекратили рытье и не возобновили работы до тех пор, пока почва не была увлажнена. Так же вели себя в садке с сухой почвой и рабочие
C. aenescens продолжали рыть, и эффективность их рытья заметно не изменилась (взвешивание не производилось, но об этом можно было приблизительно судить по размерам комков). Помещенные в тот же садок A. aberrans вначале пытались отгребать почву лапками и головой, но не переносили ее в челюстях, а вскоре прекратили рытье и не возобновили работы до тех пор, пока почва не была увлажнена. Так же вели себя в садке с сухой почвой и рабочие ![]() Formica cunicularia, собранные на Звенигородской биостанции.
Formica cunicularia, собранные на Звенигородской биостанции.
Не вызывает сомнения, что во всех, или почти всех, родах муравьев псаммофоры возникали независимо и параллельно. На это указывают как принципиальные различия в их строении у разных групп муравьев, так и то, что во многих родах мы можем проследить полный ряд развития псаммофоров. Так, например, в пределах рода Messor К. В. Арнольди (1977) выделил три последовательные стадии эволюции псаммофора. У степных (группы M. clivorum, M. rufitarsis) и горных (группы M. rufus, M. himalayanus) представителей рода имеется простой псаммофор: гулярные аммохеты короткие, лишь немного длиннее, чем у целого ряда видов Aphaenogaster, покрывают всю нижнюю поверхность головы. Некоторые пустынные виды (группы M. excursionis, M. aciculatus, M. antennatus) имеют несовершенный псаммофор, гулярные аммохеты которого также занимают практически всю нижнюю поверхность головы, но располагаются в определенном порядке. С каждой стороны имеются один — три неправильных ряда латеральных макрохет вдоль границы между нижней и боковой поверхностями головы и группа базальных гулярных макрохет в области перед затылочным отверстием. Кроме того, между макрохетами имеются дополнительные волоски, частично занимающие и переднюю часть гулярной поверхности вблизи рта. В пустынях Северной Африки и Западной и Средней Азии доминируют виды, имеющие совершенный псаммофор (в Средней Азии — M. aralocaspius, M. intermedins и M. denticulatus). У этих муравьев на нижней стороне головы имеется с каждой стороны только один-два латеральных ряда и по два-три сильно укороченных ряда длинных базальных гулярных макрохет; дополнительных волосков нет и поле передней части гулярной поверхности голое (рис:15/1/). Заключительным этапом этого ряда является специализированный псаммофор, в котором латеральные и базальные ряды макрохет сливаются и образуют один дугообразный ряд длинных гулярных аммохет, окружающих гладкую площадку на нижней стороне головы. У наших представителей Messor такой псаммофор не встречается, но он имеется у североафриканского M. caviceps и близких к нему видов. Подобные ряды можно наблюдать также в пределах родов Pogonomyrmex, Melophorus (Wheeler, 1907а), Monomorium и Chelaner.
На независимость возникновения этого признака, возможно, указывают и различия в стереотипном поведении при упаковке псаммофоров. Так, все изученные виды Cataglyphis (C. pallida, C. cinnamomea, C. foreli, ![]() C. aenescens, C. setipes) и Camponotus turcestanicus формуют комок жвалами и передними лапками, но у Cataglyphis нижнечелюстные щупики во время работы постоянно лежат на комке, а у Camponotus — прижаты к голове. Messor aralocaspius подгребает передними лапками песок с боков к голове, а затем начинает формовку вершинами передних голеней и основанием первого членика лапок и нижней стороной брюшка. Брюшко при этом постоянно прихлопывает комок снизу.
C. aenescens, C. setipes) и Camponotus turcestanicus формуют комок жвалами и передними лапками, но у Cataglyphis нижнечелюстные щупики во время работы постоянно лежат на комке, а у Camponotus — прижаты к голове. Messor aralocaspius подгребает передними лапками песок с боков к голове, а затем начинает формовку вершинами передних голеней и основанием первого членика лапок и нижней стороной брюшка. Брюшко при этом постоянно прихлопывает комок снизу.
Несложно представить, как шло формирование псаммофоров в процессе эволюции. У многих муравьев гумидных и семиаридных областей имеются отстоящие волоски на переднем крае наличника, жвалах, ментуме и нижней стороне головы. Число, расположение и длина этих волосков сильно варьируют даже у разных особей в пределах одной семьи. Так, например, у 30 случайно выбранных особей Formica lugubris, взятых из одного гнезда, число гулярных волосков на одной половине головы варьировало от 7 до 16 (среднее — 10,8; σ=2,73). Хорошо известно, что именно такие изменчивые полигенные структуры легче всего перестраиваются отбором. Можно предположить, что эти волоски помогают, хотя и незначительно, поддерживать комок почвы, выносимый из гнезда. Косвенным подтверждением этого может служить тот факт, что в Европе на песчаных почвах, где муравьи строят самые глубокие гнезда (Длусский, 1967а), обитают виды Formica с наиболее развитым отстоящим опушением нижней стороны головы (F. cinerea, F. imitans, F. pratensis, ![]() F. truncorum). Увеличение глубины гнезд при одновременном сохранении числа рабочих (что и наблюдается при переходе к жизни в аридных условиях) должно приводить к усилению отбора в этом направлении и, следовательно, к постепенному увеличению размеров и к фиксации положения щетинок.
F. truncorum). Увеличение глубины гнезд при одновременном сохранении числа рабочих (что и наблюдается при переходе к жизни в аридных условиях) должно приводить к усилению отбора в этом направлении и, следовательно, к постепенному увеличению размеров и к фиксации положения щетинок.
Как показали наши наблюдения, у Alloformica и Formica нижнечелюстные щупики принимают некоторое участие в процессе формовки комка почвы. Собственно формовка производится главным образом жвалами и передними ногами. Но по окончании работы муравьи плотно прикладывают щупики к комку, как бы контролируя его форму. При переноске комка щупики обычно свободно висят вниз, но изредка муравьи прикладывают их к комку. Несомненно, такое поведение создавало предпосылки для включения у Cataglyphis (произошедших от Alloformica) нижнечелюстных щупиков в состав псаммофора. У Camponotus, как обитающих в гумидных районах (C. herculeanus), так и у пустынных (C. turcestanicus), нижнечелюстные щупики не принимают участия в этом процессе и остаются плотно прижатыми к голове. То же наблюдается у![]() Myrmicinae, все представители которых исходно имеют укороченные щупики. Не удивительно, что у этих муравьев нижнечелюстные щупики в состав псаммофора не включаются.
Myrmicinae, все представители которых исходно имеют укороченные щупики. Не удивительно, что у этих муравьев нижнечелюстные щупики в состав псаммофора не включаются.
Легкость возникновения псаммофоров в разных группах муравьев несомненно является результатом того, что в процессе возникновения этих органов практически не нарушаются координации, сложившиеся между другими органами муравья. Во всяком случае, на начальных этапах этого процесса развитие псаммофора не требует изменения расположения мышц и связанных с ними изменений формы головы и груди.
Хотя псаммофоры описаны только у муравьев, они встречаются и у других насекомых, роющих норы и выносящих почву на поверхность. Так, например, они хорошо развиты у многих роющих ос (Sphecidae), где возникают независимо и параллельно в разных группах и зачастую достигают гораздо большей специализации, чем у муравьев.
С развитием псаммофора и повышением эффективности рытья, очевидно, связаны и изменения формы головы у некоторых пустынных видов, на которые прежде не обращали внимания. У видов с высоко развитым псаммофором мирмицинного типа часто нижняя сторона головы бывает не выпуклой, как у других муравьев, а вогнутой (рис:13/2/), что несомненно должно увеличивать емкость псаммофора. Кроме того, у многих пустынных видов параллельно возникает характерная трапециевидная форма головы с расширенным передним краем. Так, например, в роде Monomorium она возникает независимо у видов подродов Xeromyrmex и Equesimessor, с одной стороны, и южноафриканских пустынных Monomorium s. str. — с другой. Значение этого признака несложно понять, если представить себе действия рабочего, предшествующие формовке комка в псаммофоре. Вначале он отгрызает жвалами мелкие частички почвы, а затем подгребает почву под себя. При этом лапки остаются фиксированными, а муравей совершает резкие движения назад всем корпусом и одновременно головой со сложенными жвалами сверху вниз. Таким образом, передний край головы действует как мотыга. Естественно, расширение переднего края головы (как и расширение лезвия мотыги) должно повышать эффективность этого действия. Кроме того, расширение базы прикрепления жвал одновременно повышает емкость псаммофора.
У многих пустынных муравьев, в частности у палеарктических Cataglyphis и южноафриканских Ocymyrmex, ярко выражена способность рыть сухой песок, отбрасывая его под себя. Муравей при этом принимает характерную позу и очень быстро кидает песок назад передними лапками. В результате из-под него вылетает почти непрерывная струйка песка. Такое поведение очень обычно у многих одиночных ос. Среди среднеазиатских муравьев отбрасывание почвы особенно часто наблюдается у Cataglyphis pallida, часто строящего гнезда на незакрепленных или слабо закрепленных песках. В сухую погоду у входов в гнезда этого вида постоянно находится группа рабочих, очищающих таким образом вход от заплывания сухим эоловым песком. С таким поведением связано образование у C. pallida на передних лапках своеобразной «метелки» из волосков, не встречающейся у других Cataglyphis, обитающих на более плотных почвах (рис:17/4/). Сходную «метелку» мы обнаружили и у африканского псаммофильного C. bombycina (рис:17/3/). В то же время у видов, обитающих на плотных почвах, волоски на лапках утолщаются и превращаются в короткие толстые хеты (рис:17/2/).
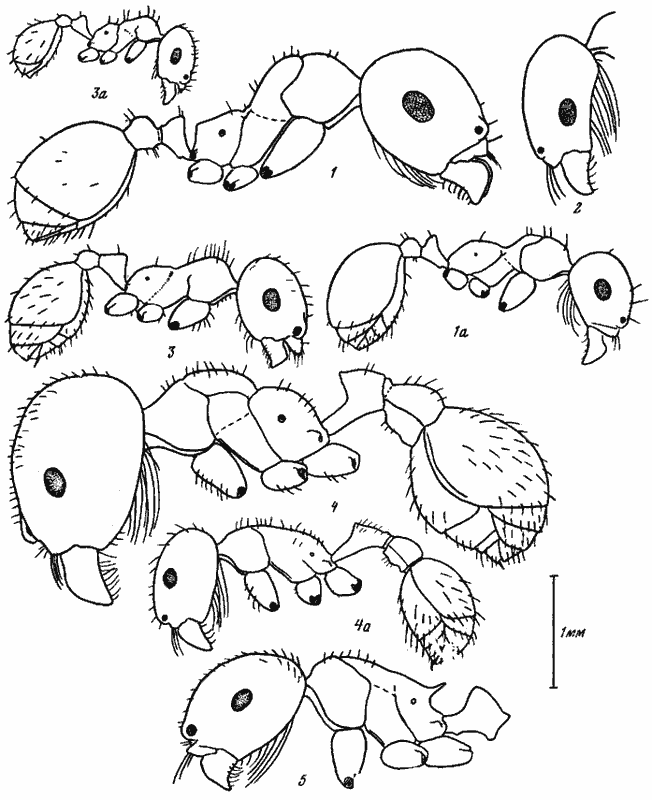
Рис. 13. Муравьи-карпофаги
1, 1a — крупный и мелкий рабочие Messor intermedins (Каракумы); 2 — голова Monomorium chobauti в профиль (Алжир); 3, 3а — Chelaner sp. (Австралия, Дениликуин); 4, 4а — Pogonomyrmex badius (США, Флорида); 5 — P. barbatus molefaciens (США, Техас)
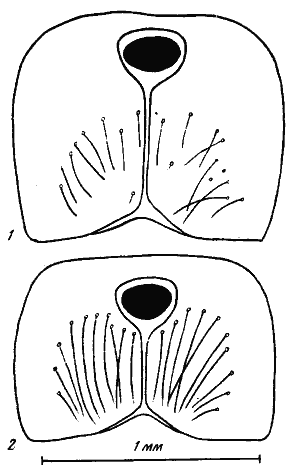
Рис. 15. Псаммофоры представителей подсемейства
1 — Messor intermedins; 2 — Pogonomyrmex barbatus molefaciens
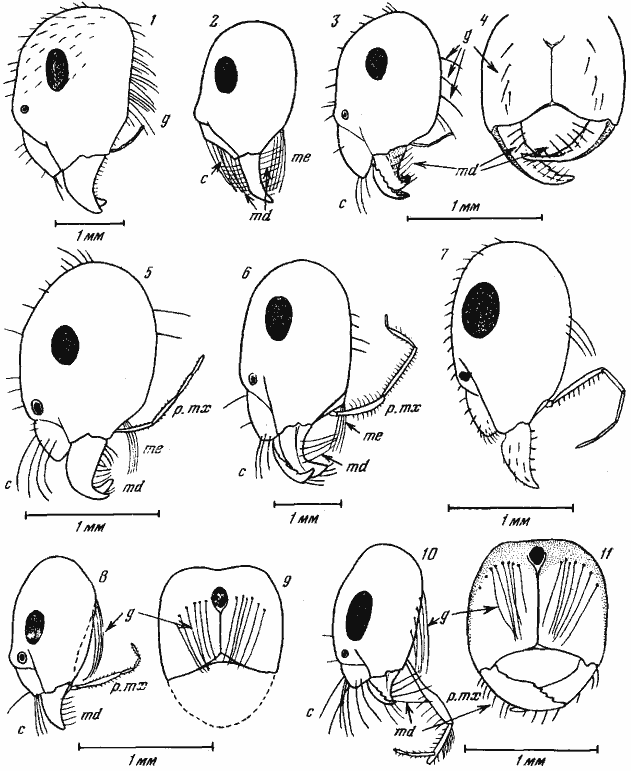
Рис. 16. Псаммофоры представителей подсемейств
2 — C. mystaceus (Калахари, по [Wheller, 1907а]);
3, 4 — Anoplolepis sp. (Африка, о-в Занзибар);
5 — Melophorus sp. А (Австралия); 6 — Cataglyphis setipes (Средняя Азия);
7 — Myrmecocystus hortideorum (Северная Америка).
10, 11 — Dorymyrmex albipes (Аргентина).
Обозначения щетинок; c — клипеальные; g — гулярные; md — мандибулярные; me — ментальные; p. mx — нижнечелюстные щупики
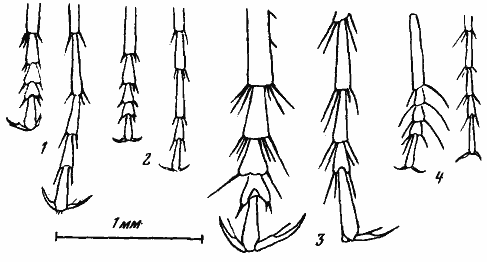
Рис. 17. Макрохеты на лапках передних (слева в каждой паре) и задних ног Cataglyphis, строящих гнезда в глинистой (1, 2) и песчаной (3, 4) почве
1 —
(лат. mandibule — челюсть, от mando — жую, грызу), первая пара челюстей у ракообразных, многоножек и насекомых. Синоним: жвала.
специальное приспособление для переноски сухой почвы, распространенн у пустынных муравьев, образован длинными отстоящими волосками (аммохетами), часто имеющими форму латинской буквы J, расположен на нижней поверхности головы.
Всего 106 видов.
Всего 60 видов.
Всего 172 вида.
Включает сокращенный Novomessor.
Всего 9 видов. Карпофаги.
Всего 48 видов.
Всего 965 видов.
Всего 23 вида.
Всего 65 видов.
Всего 30 видов.
Myrmicinae — самое большое подсемейство муравьев, включающее более 2 тыс. видов, относящихся более чем к 150 родам.
Одно из крупных подсемейств, насчитывает около 1,5 тыс. видов, относящихся к 44 родам. Широко распространены на всем земном шаре, довольно обычны в тропиках и занимают доминирующее положение в умеренных широтах.
Небольшое наиболее архаичное подсемейство муравьев. Распространено по всему земному шару, преимущественно в тропиках. Насчитывает более 230 видов.
 Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунок
Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунокгигрофил, галобионт
Старое название: Cataglyphis flavigastra
 Formica cunicularia Latreille, 1798 — прыткий степной муравей
Formica cunicularia Latreille, 1798 — прыткий степной муравейнеобратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.
Старое название: Messor rufitarsis
ксеробионт
гигрофил
Старое название: Cataglyphis cinnamomea
ксеробионт
ксеробионт
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
 Formica truncorum Fabricius, 1804 — красноголовый муравей
Formica truncorum Fabricius, 1804 — красноголовый муравейВсего 4 вида.
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
сборное название насекомых подотряда жалящих перепончатокрылых (кроме пчел и муравьев). Длина до 40 мм (шершни). Ок. 23 тыс. видов. Распространены широко. Опылители растений. Личинки некоторых ос паразиты других насекомых.
Старое название: Cataglyphis bombycina
организм, питающийся семенами растений.
«медовые муравьи»
Всего 22 вида.
Старsые названия: M. hortideorum, M. hortusdeorum
Старое название: Araucomyrmex tener






