






Пустыня как среда обитания муравьев
Животные и растения пустынь издавна привлекали внимание исследователей. Хотя в последние годы число работ по изучению пустынь резко возросло, в первую очередь в связи с необходимостью хозяйственного освоения аридных районов, работы биологов носят по преимуществу статичный характер: изучаются продуктивность различных пустынных биогеоценозов, пути повышения их продуктивности, морфофизиологические приспособления, позволяющие животным и растениям выживать в этих суровых условиях. Эволюционный подход при этом, как правило, отсутствует. В то же время лишь эволюционный подход позволяет понять, почему возникают те, а не иные приспособления или та, а не иная структура биогеоценоза. Одновременно изучение путей адаптации организмов к жизни в пустыне может дать богатейший материал для изучения общих закономерностей филогенеза. Не случайно классическими примерами конвергенции являются, с одной стороны, водные организмы (акулы, ихтиозавры, китообразные, скаты и камбалы), а с другой — животные и растения аридных областей (тушканчики Евразии и кенгуровые крысы Америки; цокоры Азии, гоферы Северной Америки и тукотуко Южной Америки; слепушонки Евразии и Bathyergidae Африки; кактусы Америки и молочаи Африки и т. д.). Как справедливо отмечал Д. Н. Кашкаров (Кашкаров, Коровин, 1936, с. 34), «в климате пустыни имеется ряд моментов, противоречащих основным требованиям организмов. Последние должны так или иначе, изменяя свою структуру, или поведение, или физиологические особенности, приспособляться к климатическим условиям пустыни... И в этом направлении в пустыне действует самый жестокий естественный отбор».
Основными, общими для всех пустынь Земли, лимитирующими факторами являются следующие:
1. Недостаток влаги. В пустынях выпадает менее 250 мм осадков в год, причем распределение их неравномерно. В некоторых пустынях их практически не бывает: в Центральной Сахаре (Эль-Минья, Асьют, Вади-Хальфа) в среднем выпадает 0,1 мм осадков в год, в Атакаме (Икике) — 3 мм/год, Кашгарии (Черчен) —9 мм/год (Петров, 1973).
2. Неблагоприятные температурные условия. Самые высокие на Земле температуры воздуха были зарегистрированы в пустынях: Долина Смерти в Калифорнии — +54,7°, Ливийская пустыня (Азизи) — +58°, Каракумы (Репетек) — +49,9°. Еще сильнее нагревается поверхность почвы. Самые высокие температуры были отмечены в пустынях на побережье Красного моря (83,5°), в Сахаре (81°) и в Каракумах (Репетек, 79,4°). В континентальных пустынях умеренного пояса отмечается огромный перепад минимальных и максимальных температур, доходящий до 120° (Репетек — от -40 до +79,4°) (Петров, 1973). В холодных высокогорных пустынях годовая амплитуда температуры поверхности почвы также достигает 100° (на Памирской биостанции от -45,7 до +56,8°), но максимальные температуры гораздо ниже (в Памирской пустыне температура воздуха летом +20—22°, температура поверхности почвы — до 56,8°) (Вальтер, 1975). Суточные перепады температур также велики и в пустынях Средней Азии составляют 20—30°.
3. Скудость и сезонная неравномерность пищевых ресурсов животных является следствием неблагоприятного климата. Показателем обилия ресурсов является продукция фитомассы. Так, фитомасса лесов умеренной зоны составляет 150—500 т/га, типчаково-ковыльных степей — 18—34 т/га, саксауловых лесов Средней Азии — 55 т/га, эфемерово-полынных пустынь — 11—14 т/га, полынных пустынь — 4,6 т/га, солянковых пустынь — 2—12,5 т/га, такыров — 0,1—0,6 т/га (Родин, 1961). В пустынях Западной Африки (осадки 100—250 мм/год) годовая продукция сухого вещества составляет 0,7—1,6 т/га, а в полупустынях (350—600 мм/год) — 3,2—5,5 т/га. Даже во влажные годы в наиболее богатых биоценозах Центральных Каракумов проективная полнота растительности не превышает 45% (Нечаева, 1970), а в экстрааридных и некоторых типах аридных пустынь (бедлендовые, такыровые) высшая растительность практически отсутствует (биомасса 0,001—0,01 т/га) (Петров, 1973).
Количество пищи сильно меняется в пустынях и в течение года. Вегетация растений приурочена к влажному сезону. В пустынях Средней Азии весной почва покрывается эфемерами, которые летом полностью высыхают и развеиваются ветром. Так, в Центральных Каракумах развитие эфемеровой растительности в годы с незначительным (90—110 мм) количеством осадков заканчивается в 1—2-й декадах мая, а в более влажные годы (130—160 мм осадков) — в 1—2-й декадах июня (Антонова, 1970). По нашим данным (Длусский, Сапарлыев, 1975), в саксауловых лесах дельты Мургаба основная продукция семян приходится на апрель (65%) и май (23%).
В литературе существует множество определений понятия «пустыня». Не вдаваясь в обсуждение этого вопроса, отмечу, что я придерживаюсь определения М. П. Петрова (1973, с. 8), согласно которому под пустынями понимаются «территории с предельно засушливым климатом (осадков менее 200—250 мм/год, они нерегулярны и испарение преобладает над ними), иногда с отмершей речной сетью или с редкими транзитными реками, без постоянного поверхностного стока, с засоленными почвогрунтами и солевыми корами, с господством сульфидов и хлоридов. Это территории, где органическая жизнь подавлена, растительный покров и животный мир очень скудны, а в почвах миграции солевых растворов преобладают над биогенными процессами и где земледелие возможно только при орошении».
Я не хотел бы также вступать в полемику по поводу термина «полусаванна», предложенному П. Н. Овчинниковым (1940) для обозначения своеобразных подгорных и предгорных эфемеровых ландшафтов юга Средней Азии (Копетдаг, Бадхыз, Западный Тянь-Шань, Гиссаро-Дарваз) и в настоящее время довольно распространенному среди геоботаников (Коровин, 1958; Лавренко, 1962; и др.). Я полностью согласен с О. Л. Крыжановским (1965), что термин неудачен, поскольку эти ландшафты к настоящей саванне не имеют никакого отношения и скорее являются переходом от пустыни к низкогорным степям. И все же я использую термин «полусаванна», поскольку, во-первых, своеобразие этих ландшафтов несомненно, а во-вторых, неудачность устоявшегося термина, как мне кажется, не является достаточным основанием для переделки терминологии (пользуемся же мы такими неудачными терминами, как «индеец», «кровное родство» или «новолуние»).
Пустыни располагаются в трех климатических поясах (умеренном, субтропическом и тропическом) и по подсчетам разных авторов занимают 29—32 млн. км2, или 19—23% поверхности суши (Петров, 1973). В настоящее время на Земле имеются 5 крупных пустынных областей (рис:1): Великий Пояс пустынь Старого Света (или пустыни древнего Средиземья), пустыни Мексики и юга США, пустыни Южной Америки, пустыни Южной Африки и пустыни Австралии. В дальнейшем общая характеристика дается по работам Мейгса (Meigs, 1953), T. В. Власовой (1966), М. П. Петрова (1973), Г. Вальтера (1975), биогеографическое районирование — по А. Г. Воронову (1963) и О. Л. Крыжановскому (1965).

Рис. 1. Пустыни и семиаридные области Земли
а — пустыни (кроме полярных);
б — семиаридные области.
Африка: 1 — Сахара; 2 — Ливийская пустыня; 3 — Нубийская пустыня; 5 — Намиб; 6 — Калахари; 7 — Большое Кару.
Азия: 4 — Синайская пустыня; 8 — Сирийская пустыня; 9 — Большой Нефуд; 10 — Малый Нефуд; 11 — Руб-эль-Хали; 12 — Деште-Ке-вир; 13 — Деште-Лут; 14 — Тар; 15 — Устюрт; 16 — Каракумы; 17 — Кызылкумы; 18 — Бетпак-Дала; 19 — Муюнхум; 20 — Такла-Макан; 21 — Гоби; 22 — Алашань; 23 — Цайдам; 24 — высокогорные пустыни Тибета, Каракорума и Памира.
Австралия: 25 — Большая Песчаная пустыня; 26 — пустыня Гибсона; 27 — Большая Пустыня Виктория; 28 — пустыня Симпсона.
Северная Америка: 29 — Большой Бассейн; 30 — Мохаве; 31 — Сонорская пустыня; 32 — Чиуауа.
Южная Америка; 33 — прибрежные пустыни Чили (в том числе Атакама) и Перу и высокогорные пустыни на западных склонах Анд; 34 — пустыни на восточных высокогорных плато Анд; 35 — пустыни Патагонского плато.
Великий Пояс пустынь Старого Света простирается от атлантического побережья Африки (17° з.д.) до Центрального Китая (112° в.д.) и захватывает Северную Африку, значительную часть Азии (около 35% территории) и крайний юго-восток Европы (около 1% территории). В настоящее время он подразделен на 5 частично изолированных областей, между которыми, однако, может происходить некоторый обмен флорами и фаунами. Западную часть Пояса составляет Сахара, крупнейшая пустыня мира, покрывающая площадь около 7 млн. км2. Северная часть ее лежит в зоне субтропического, центральная и южная — в зоне тропического климата. Между Нилом и Красным морем располагаются Аравийская и Нубийская пустыни. Пустынями занят почти весь Аравийский полуостров. Горы Загрос отделяют пустыни Аравийского полуострова и Месопотамии, фаунистически очень сходные с Сахарой, от пустынь Иранского нагорья, лежащих в субтропическом поясе. На севере Туркмено-Хорасанские горы отделяют Иранское нагорье от пустынь Средней Азии и Казахстана, лежащих в умеренном и субтропическом поясах. Этот барьер не является непреодолимым: фаунистический обмен может осуществляться как по долине Теджена (Герируд), так и через невысокие горы с ксерофитной полусаванной растительностью (Копетдаг). На юго-востоке узкая полоса саванн отделяет пустыни Иранского нагорья от тропических пустынь Индостана (Тар, Тал). По дюнам вдоль побережья Персидского и Оманского заливов и Аравийского моря может осуществляться связь между пустынями Индостана и Аравийского полуострова. Через Кашгарские ворота и долину Черного Иртыша пустыни Казахстана соединяются с Джунгарией, входящей в состав пустынь Центральной Азии, расположенных на территории МНР и КНР. На плоскогорьях Тибета и Памира лежат своеобразные холодные высокогорные пустыни.
Пустыни Казахстана и Средней Азии, расположенные на территории СССР, представляют фактически одно целое и делятся лишь условно. Обычно выделяют следующие крупные области:
Пустыни Прикаспийской низменности вдоль северного побережья Каспийского моря;
Мангышлак на полуострове Мангышлак;
Устюрт между Аральским и Каспийским морем;
Каракумы между Туркмено-Хорасанскими горами (Копетдаг, Парапомиз), долиной Амударьи и плато Устюрт;
Кызылкумы, ограниченные долинами Амударьи и Сырдарьи, Аральским морем и долиной Зеравшана и отрогами Алая;
Каршинская «степь» к югу от Кызылкумов между долинами Амударьи, Зеравшана и отрогами Алая.
Приаральские Каракумы и Барсуки к северу и северо-востоку от Аральского моря.
Бетпак-Дала к северу от долины Чу.
Муюнкумы между долиной Чу и отрогами Тянь-Шаня (Каратау, Киргизский хребет).
Прибалхашские пустыни (Таукум, Сарытаукум, Сары-Ишикотрау, Прибалхашские Каракумы и др.) к югу и востоку от оз. Балхаш.
Пустыни Великого Пояса относятся к Сахаро-Гобийской пустынной зоогеографической подобласти области Древнего Средиземья Голарктики.
На юге Африки пустыни Калахари и Намиб лежат в тропическом поясе, а Большое Кару — в субтропическом. В пустыне Намиб, вытянутой полосой шириной 100—150 км вдоль атлантического побережья Африки, выпадает всего 15—43 мм осадков в год, однако близость океана обусловливает здесь высокую (до 70—80%) относительную влажность воздуха. Часть этой влаги конденсируется на поверхности почвы, благодаря чему растительность здесь местами довольно обильна. В Калахари выпадает до 154 мм осадков в год, а в большом Кару — до 230 мм., Между этими тремя пустынями может осуществляться частичный обмен флорами и фаунами через разделяющие их полупустыни и саванны. Эти пустыни относятся к Южноафриканской подобласти Эфиопской зоогеографической области.
Пустыни Северной Америки простираются между 22 и 44° северной широты и занимают юго-запад США и значительную часть территории Мексики. Пустыни Большого Бассейна (штаты Орегон, Айдахо, Невада, Юта, Аризона, США) лежат в зоне умеренного пояса и являются климатическим аналогом пустынь Средней Азии. Пустыня Мохаве, также лежащая в умеренном поясе, отличается очень сухим и жарким климатом. В субтропическом и тропическом поясах лежат Сонорская пустыня в широком смысле (п-ов Калифорния и штат Сонора, Мексика, и юго-запад штата Аризона, США) и пустыня Чиуауа (штаты Чиуауа и Дуранго, Мексика, и юго-запад штата Нью-Мексико, США). Все пустыни Северной Америки, как и пустыни Средней Азии и Казахстана, представляют собой единое целое и границы между ними условны. Все они относятся к Сонорской зоогеографической области Голарктики.
В Южной Америке пустыни занимают узкую полосу шириной до 150 км вдоль побережья Тихого океана в пределах Чили и Перу между 7 и 31° ю.ш. и участок на Патагонском плато между реками Рио-Колорадо и Санта-Крус. Прибрежная пустыня отличается, как и все приокеанические пустыни, очень малым количеством осадков, но местами сравнительно высокой влажностью воздуха. В Атакаме (участок между 22 и 27° ю.ш.) и некоторых районах Перу высшая растительность практически отсутствует. Пустыни Патагонского плато обликом напоминают памирские высокогорные пустыни: растительность здесь представлена ксерофитными подушечниками, что является результатом частых и сильных ветров. Однако климат этих пустынь значительно теплее памирских. Прибрежная пустыня и Патагонское плато разделены Андами, но относятся к одной Чилийско-Патагонской подобласти Неотропической зоогеографической области. Общая площадь этих пустынь невелика — всего 1,8 млн. км2, или около 10% территории.
В Австралии пустыни занимают всю центральную часть материка и общая их площадь составляет 3,4 млн. км2, или 49% территории. Все они фактически представляют одно целое и носят поэтическое название «Красное сердце Австралии». В основном эти пустыни лежат в тропическом поясе и лишь равнина Налларбор — в субтропическом. Все пустыни Австралии относятся к Новоголландской подобласти Австралийской зоогеографической области.
Хотя следы наличия пустынных областей в виде отложений ископаемых солей известны по крайней мере с кембрия (Lotze, 1938), а растения-ксерофиты найдены в верхнем палеозое (Марков, 1956), формирование пустынь современного облика началось в верхнем мелу. В это время в северном полушарии приблизительно по 55° с. ш. проходила граница между Гренландской и Геленденской ботанико-географическими провинциями, причем в первой в значительной степени были представлены листопадные субтропические элементы (Криштофович, 1955). В тропической Геленденской провинции в это же время сформировалась субаридная область (500—800 мм осадков в год), включавшая юг Руссоскандии (территория современной Центральной Европы), северо-восток Лемурии (современная Аравия), Иранию и центральную часть Палеазии (современная территория Казахстана, Средней и Центральной Азии) (Синицын, 1967).
В течение палеоцена, эоцена и олигоцена на базе Гренландской флористической провинции сформировалась Тургайская флористическая область, а на базе Геленденской провинции — Полтавская флористическая область, причем граница между ними постепенно сдвигалась к югу (Криштофович, 1955). Представление о ксерофитной полтавской флоре эоцена дают находки близ оз. Ер-Ойландуз (Южная Туркмения). Здесь обнаружены ископаемые представители 20 семейств растений, причем в основном это были вечнозеленые деревья и кустарники с мелкими кожистыми листьями, с морщинистой поверхностью и выступающими жилками. В современной флоре древнейшими представителями полтавской ксерофитной флоры являются Ephedra пустынь Центральной и Средней Азии и Северной Америки и Welwitschia пустыни Намиб (Криштофович, 1955). Помимо аридных областей Великого Пояса и Южной Европы, засушливые области в палеогене существовали на территории США, Перу, Колумбии, южноафриканских пустынь и пустынь Центральной Австралии, причем, как и сейчас, эти области отделялись друг от друга морем или обширными гумидными областями (Шварцбах, 1955).
В миоцене в связи с дальнейшим похолоданием и иссушением климата на территории современного Великого Пояса пустынь и пустынь Северной Америки формируются травянистые равнины. Растительность их складывалась автохтонно на базе полтавской и тургайской флор путем почти полной элиминации древесных и кустарниковых элементов и бурного родо- и видообразования травянистых форм (Криштофович, 1955). На территории современных Кызылкумов выпадало 200—300 мм осадков в год (Синицын, 1967). В конце плиоцена сформировались настоящие пустыни современного облика, территории которых приблизительно соответствовали территориям современных пустынь (Марков, 1956). На территории современных Кызылкумов в плиоцене выпадало, как и сейчас, менее 200 мм осадков в год (Синицын, 1967). В течение четвертичного периода границы пустынь подвергались довольно значительным колебаниям. В ледниковые эпохи аридные зоны сужались, а в межледниковья — расширялись (Берг, 1947). Значительно более влажным в плейстоцене был климат Сахары и Большого Бассейна Северной Америки (Шверцбах, 1955; Петров, 1973). Однако в общих чертах положение аридных областей в четвертичном периоде соответствовало современному.
Таким образом, палеоклиматологические данные указывают, что пять основных аридных областей всегда были разделены океанами и территориями с гумидным климатом, так что их биогеоценозы должны были формироваться независимо. На это указывает и высокий эндемизм пустынных флор и фаун. Даже если сравнить флоры и фауны пустынь Старого Света и Северной Америки, сформировавшиеся на основе голарктической флоры и фауны, мы увидим, что семейства млекопитающих или роды растений, характерные только для Неоарктики или Палеарктики, — это, как правило, семейства или роды, сформировавшиеся и обитающие в аридных областях. Так, в Неоарктике не встречаются такие характерные растения пустынь Азии, как Haloxylon, Calligonum, Salsola, муравьи Messor и Cataglyphis, грызуны из семейств Spalaeidae и Dipodidae, лошади, ослы, верблюды и т. д. В то же время, в тех таксонах, отдельные представители которых обитают и в пустынях Палеарктики и в пустынях Неоарктики, имеется много видов, обитающих в гумидных областях Голарктики (растения Artemisia, Festuca, муравьи Formica, грызуны из семейств Sciuridae и Muridae).
Некоторый обмен флорами и фаунами происходил между пустынями Северной и Южной Америки и Северной и Южной Африки. На это указывает биполярное распространение ряда аридных животных и растений. Так, распространены в Новом Свете некоторые роды кактусов, муравьи Pogonomyrmex. Жуки Julodis, Adesmia, Pentodon, муравьи Messor и некоторые растения встречаются в Северной и Южной Африке, но отсутствуют в Экваториальной. Л. С. Берг (1947) связывает биполярное распределение африканских животных с иссушением климата в плейстоцене, причем обращает внимание на то, что распространение шло с севера на юг. Движение на юг скорее всего происходило по прибрежным дюнам. На это указывает наличие большого количества североафриканских элементов в песчаных пустынях и высокий эндемизм каменистых пустынь Южной Африки (Петров, 1973) и общность некоторых растений (например, Aristida) песчаных пустынь и интразональных прибрежных дюн Африки (Вальтер, 1975).
Вопрос об истории формирования фаун и флор отдельных пустынных областей Великого Пояса, в частности пустынь Казахстана и Средней Азии, довольно запутан, и в литературе встречается множество представлений, зачастую диаметрально противоположных. Обстоятельный обзор взглядов на происхождение фауны Средней Азии дается в монографии О. Л. Крыжановского (1965). Анализ фауны муравьев аридных областей Палеарктики, который будет приведен ниже, полностью подтверждает точку зрения этого автора, согласно которой фауна Средней Азии формировалась в значительной мере автохтонно, но в то же время постоянно обогащалась пришлыми элементами. В свою очередь большинство мигрантов уже на территории Средней Азии прошли путь автохтонной переработки и образовали эндемичные виды. Аналогичные процессы происходили и в других частично изолированных областях Великого Пояса, так что вся его фауна может рассматриваться как единое целое.
Таким образом, пустыни Земли представляют собой результат уникального эволюционного эксперимента природы. В пяти изолированных областях с одинаковыми ландшафтами и климатом (и соответственно сходными векторами отбора) на фоне очень жесткого отбора на базе различных фаун возникли аналогичные биогеоценозы. Сравнение этих областей может дать богатейший материал для изучения закономерностей эволюционного процесса, в частности для изучения общих закономерностей адаптации, проблем преадаптации и коадаптации, проблемы направленности эволюционного процесса.
Удобной моделью для таких исследований являются муравьи. Прежде всего это обусловливается обилием этих насекомых во всех пустынях земного шара, причем удельный вес муравьев среди других беспозвоночных увеличивается по мере увеличения аридности климата. Так, например, в прериях США продукция муравьев составляет 1—14%, в полупустынях — 7—9% и в пустынях — 11—15%. В отдельных пустынях муравьи составляют 96—99,7% от всех беспозвоночных (Petal, 1978). Во-вторых, муравьи представляют собой довольно компактную группу (семейство отряда Hymenoptera). В-третьих, муравьи практически не имеют конкурентов среди других животных и поэтому эволюировали как в значительной мере автономный комплекс. Это позволяет, изучая эволюцию муравьев как таксономической группы, изучать также и эволюцию одного из коадаптивных комплексов. И, наконец, в-четвертых, муравьи обладают весьма сложным и разнообразным поведением, что позволяет изучать не только эволюцию морфофизиологических адаптации, но и эволюцию поведения.
Задачей работы было изучить, по возможности полно, весь комплекс адаптации, позволяющих муравьям существовать в условиях пустыни. При этом представлялось целесообразным рассмотреть не только приспособления, возникшие у муравьев в связи с жизнью в пустынях (как это обычно делается), но и приспособления, возникшие в гумидных условиях, но позволяющие этим насекомым освоить пустыни.
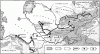
Рис. 2. Карта района исследований
1 — территории на высоте более 2000 м;
2 — территории на высоте 500—2000 м (на территории СССР это, как правило, полусаванны, подгорные полупустыни, сухие степи и степи);
3 — пустыни (на территории СССР);
4 — автомобильные маршруты с остановками для фаунистических сборов;
5 — точки, где была довольно полно собрана фауна и сделаны количественные учеты муравьев;
6 — точки стационарных исследований: заповедник «Аксу-Джабаглы» — 1958 г.; Кызылкумская пустынная станция Института ботаники АН УзССР (Аяк-Гужумды, Кульджуктау, Дарья-сай) — 1961, 1965 гг.; пос. Артезианский — 1961, 1963 гг.; Акыбай — 1969, 1970, 1971 гг.; Центрально-Каракумский стационар Института пустынь, АН ТССР (Каррыкуль) — 1970 г.; Карши — 1974 г.; Варганза — 1978 г.
Полевые работы велись тремя способами:
1. Автомобильные маршруты с краткими остановками для сбора фауны.
2. Более длительные (1—5 дней) остановки, во время которых не только собиралась фауна, но и делались количественные учеты муравьев.
3. Стационарные исследования (более недели), во время которых делались количественные учеты и изучалась биология муравьев.
Стационарные исследования проводились в следующих точках:
1. Кызылкумская пустынная станция Института ботаники АН УзССР (Аяк-Гужумды, Кульджуктау, Дарья-сай). Расположена в Юго-Западных Кызылкумах в предгорьях останцовых гор Кульджуктау. Описание природных условий дано в работах И. Ф. Момотова (1961) и X. В. Хамзина (1963).
2. Карак-Атинская впадина (Карак-Ата, пос. Артезианский), Юго-Западные Кызылкумы.
3. Центрально-Каракумский стационар Института пустынь АН ТССР (Каррыкуль). Описание дано в монографии «Растительность центральных Каракумов и ее продуктивность» (1970).
4. Акыбайское лесничество Марыйского лесхоза ТССР (Акыбай), расположенное в высохшей дельте Мургаба к северу от Мары. Описание в статье Н. П. Кривошеиной и А. А. Захарова (1975).
5. Стационар Каршинской экспедиции Института медицинской паразитологии и тропической медицины, расположенный в 20 км к западу от г. Карши (Карши).
6. Окрестности с. Варганза Кашкадарьинской области УзССР, верховья р. Кашка-дарья.
7. Бадхызский заповедник (Бадхыз).
Маршруты, точки, в которых делались учеты муравьев, и районы стационарных исследований показаны на рис:2.
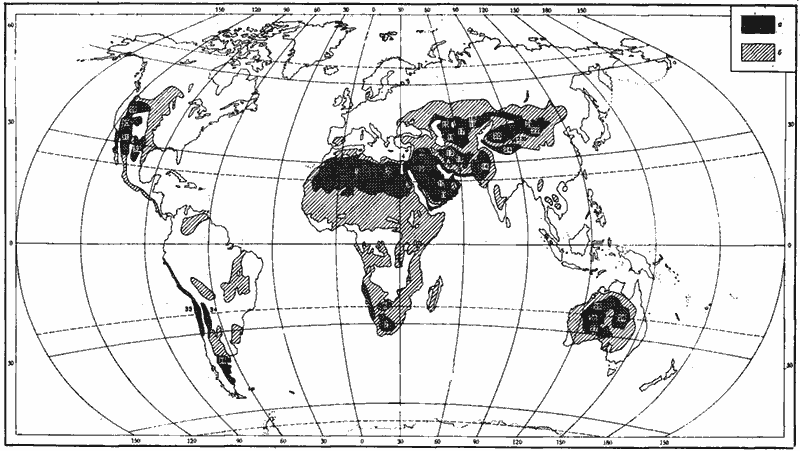
Рис. 1. Пустыни и семиаридные области Земли
а — пустыни (кроме полярных);
б — семиаридные области.
Африка: 1 — Сахара; 2 — Ливийская пустыня; 3 — Нубийская пустыня; 5 — Намиб; 6 — Калахари; 7 — Большое Кару.
Азия: 4 — Синайская пустыня; 8 — Сирийская пустыня; 9 — Большой Нефуд; 10 — Малый Нефуд; 11 — Руб-эль-Хали; 12 — Деште-Ке-вир; 13 — Деште-Лут; 14 — Тар; 15 — Устюрт; 16 — Каракумы; 17 — Кызылкумы; 18 — Бетпак-Дала; 19 — Муюнхум; 20 — Такла-Макан; 21 — Гоби; 22 — Алашань; 23 — Цайдам; 24 — высокогорные пустыни Тибета, Каракорума и Памира.
Австралия: 25 — Большая Песчаная пустыня; 26 — пустыня Гибсона; 27 — Большая Пустыня Виктория; 28 — пустыня Симпсона.
Северная Америка: 29 — Большой Бассейн; 30 — Мохаве; 31 — Сонорская пустыня; 32 — Чиуауа.
Южная Америка; 33 — прибрежные пустыни Чили (в том числе Атакама) и Перу и высокогорные пустыни на западных склонах Анд; 34 — пустыни на восточных высокогорных плато Анд; 35 — пустыни Патагонского плато.
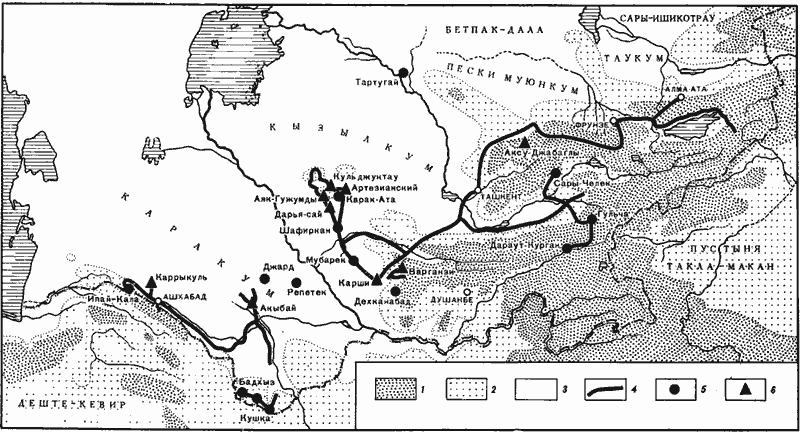
Рис. 2. Карта района исследований
1 — территории на высоте более 2000 м;
2 — территории на высоте 500—2000 м (на территории СССР это, как правило, полусаванны, подгорные полупустыни, сухие степи и степи);
3 — пустыни (на территории СССР);
4 — автомобильные маршруты с остановками для фаунистических сборов;
5 — точки, где была довольно полно собрана фауна и сделаны количественные учеты муравьев;
6 — точки стационарных исследований: заповедник «Аксу-Джабаглы» — 1958 г.; Кызылкумская пустынная станция Института ботаники АН УзССР (Аяк-Гужумды, Кульджуктау, Дарья-сай) — 1961, 1965 гг.; пос. Артезианский — 1961, 1963 гг.; Акыбай — 1969, 1970, 1971 гг.; Центрально-Каракумский стационар Института пустынь, АН ТССР (Каррыкуль) — 1970 г.; Карши — 1974 г.; Варганза — 1978 г.
эволюционно сложившаяся, пространственно ограниченная, длительно самоподдерживающаяся, однородная экологическая система, в которой функционально взаимосвязаны живые организмы и окружающая их абиотическая среда.
необратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.
общая масса особей одного вида, группы видов или сообщества в целом на единицу поверхности или объема местообитания. Биомасса растений называется фитомассой, биомасса животных зоомассой.
Всего 106 видов.
Всего 65 видов.
Всего 60 видов.
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
животные, не имеющие позвоночника: простейшие, губки, кишечнополостные, черви, моллюски, членистоногие, иглокожие и др.; примерно 3/4 видов всех животных.






