






Вопросу о полиморфизме муравьев посвящена обширная литература (обзор: Wilson, 1971). С разной степенью полноты в настоящее время изучены такие вопросы, как классификация типов полиморфизма, феноменологическая сторона его эволюции (т.е. построение рядов без функционального обоснования), разделение функций (главным образом у видов с развитым полиморфизмом), механизмы дифференциации в онтогенезе и регуляция соотношения каст в семье, регуляция соотношения каст у видов с развитым полиморфизмом в процессе эволюции. В то же время вопрос о начальных причинах и путях становления полиморфизма рабочей касты у общественных насекомых до сих пор даже не ставился.
В связи с различными толкованиями термина «полиморфизм» необходимо коротко остановиться на вопросах терминологии. Первоначально под полиморфизмом у общественных насекомых (как и под полиморфизмом вообще) понималось наличие нескольких четко отличающихся форм одного пола (Wheeler, 1908с). В последние годы получило распространение определение Э. Вильсона, опубликовавшего серию работ, посвященных полиморфизму у муравьев (Wilson, 1953, 1954, 1968, 1971). Согласно его определению, «полиморфизм — это случай неизометрического относительного роста, наблюдающийся при значительной вариации в размерах в нормальной взрослой колонии, при котором производятся особи с различными пропорциями в крайних размерных группах». Соответственно при мономорфизме «рабочие нормальной взрослой колонии либо изометричны, либо обнаруживают ограниченную вариабельность в размерах, либо то и другое; их частотное распределение унимодально» (Wilson, 1971, с. 140). Таким образом в качестве основного признака полиморфизма принимается не наличие обособленных групп (би- или полимодальность распределения), а изменение пропорций, связанное с изменением размеров (аллометрия). Последовательное применение этого критерия породило такой нелепый термин, как «двуфазный мономорфизм», обозначающий наличие двух четко обособленных форм, пропорции которых сходны (Gray, 1973).
Поскольку причиной простой аллометрии является неравномерность темпов роста разных частей тела, изометрия может рассматриваться лишь как редкий и частный случай аллометрии, при котором темпы роста сравниваемых частей близки (Мина, Клевезаль, 1976). В ряде случаев выявление аллометрии зависит от метода обработки данных. Так, классическими методами у Messor semirufus была обнаружена идеальная изометрия (т.е. мономорфизм в понимании Вильсона), а методом построения полиноминальных регрессий с помощью ЭВМ — трехфазная аллометрия (т.е. довольно совершенный полиморфизм в понимании Вильсона) (Baroni Urbani, 1976). Простая (монофазная) аллометрия обычно не является результатом эволюции. Это можно легко понять, сравнив два растения или двух животных одного вида, выросших при недостатке и изобилии пищи. Так, например, в потомстве одной линейной самки Drosophila melanogaster (линия ND-32), выросшем при разном обеспечении кормом, по нашим измерениям, длина тела самцов (n=66) варьировала от 1,88 до 2,55 мм, причем зависимость длины крыла от длины тела лучше всего описывается уравнением у=0,70x0,835. У крупных особей (2,37—2,55 мм, n=28) крыло в 1,66 раз короче тела, а у мелких (1,88—2,10 мм, n=15) — в 1,63 раза. Различие это статистически достоверно (σ1=0,0094; σ2=0,0034; t=2,85; Р<0,01).
В то же время бимодальность распределения указывает на наличие в семье механизмов, препятствующих появлению особей среднего размера, что несомненно является результатом эволюции. Равным образом результатом эволюции является и нарушение правильности аллометрии. Полифазная аллометрия явно свидетельствует о появлении нескольких каналов онтогенеза, аналогичных классическим адаптивным модификациям (например, воздушные и водные листья растений-амфибионтов) /В настоящее время можно считать доказанным, что касты у муравьев определяются не генетически, а трофически или гормонально, так что разные формы самок этих насекомых (самки, рабочие, солдаты) фактически являются адаптивными модификациями./. В связи с этим кажется целесообразным вернуться к исходному значению термина и говорить о полиморфности рабочей касты муравьев лишь в том случае, когда наблюдается отчетливое бимодальное (или полимодальное) частотное распределение по размерам в пределах одной семьи.
На основе анализа соотношения мономорфных и полиморфных видов 43 локальных фаун Э. Вильсон (Wilson, 1968) приходит к выводу, что основным фактором, определяющим это соотношение, является географическая широта, отмечая при этом, что в пустынях процент полиморфных видов несколько увеличивается. Правда, при этом остается не совсем ясным, каким критерием пользовался автор для разделения полиморфных и мономорфных видов. Так, для Средней Азии («Туркестан») указываются 7 полиморфных видов из 39 (17,9%). В Каракумах из 34 зарегистрированных видов у половины (17) наблюдается хорошо выраженная аллометрия. По данным же Вильсона, самый высокий процент полиморфных видов наблюдается на Ямайке (41,2%) и в тропиках Бразилии (38%). Если же говорить о полном диморфизме, то он наблюдается только у одного среднеазиатского вида — Pheidole pallidula.
Приведенная выше работа Э. Вильсона — единственная попытка сравнительного анализа степени вариабельности видов в разных фаунах муравьев. До сих пор никто не обращал внимания на тот факт, что в гумидных областях Палеарктики у большинства видов изменчивость рабочей касты гораздо ниже, чем в аридных. В качестве иллюстрации можно привести данные табл.17, где проводится сравнение вариабельности муравьев шести локальных фаун СССР, расположенных в лесной (Звенигородская биостанция МГУ, Московская область; Уссурийский заповедник, Приморский край), степной (Кургальджинский заповедник, Казахская ССР) и пустынной (урочище Тартугай, Северные Кызылкумы, Казахская ССР; урочище Каррыкуль, Центральные Каракумы, Туркменская ССР; Акыбайский участок Марыйской области, ТССР) зонах. В качестве индекса вариабельности (IV) взято отношение максимальной длины груди к минимальной. Из таблицы видно, что в лесной зоне больше половины видов почти мономорфны и не вариабельны (IV=1,00—1,25). В пустынях такие виды составляют всего 30—45%. В то же время у значительной части пустынных видов (44—72%) длина груди крупных рабочих более чем в 1,5 раза больше, чем у мелких, тогда как в лесной и степной зонах такие виды составляют 25—32%. В среднем индекс вариабельности у лесных и степных видов был 1,32—1,33, а у пустынных — 1,50—1,61. Хотя вес и размер добычи, которую приносит в гнездо один и тот же муравей, может очень сильно варьировать, для каждой особи существуют некоторые верхние и нижние границы и оптимальный вес груза. Естественно, эти параметры будут связаны с размерами особей. Так, муравьи-листорезы несут в гнездо куски листьев, размер которых тесно скоррелирован (r=+0,8) с размером муравья (Lutz, 1929). Такая же связь отмечена для Formica, Messor (Новиков, 1929) и Dorylinae (Schneirla, 1971). Летом 1975 г. на территории Звенигородской биостанции МГУ на дорожках гнезд ![]() Formica rufa было собрано 170 муравьев, несущих грузы. У каждого муравья измерялась длина груди, а груз сразу же взвешивался на торзионных весах. Результаты измерений показаны на графиках рис:36. В целом корреляция между размером насекомого и весом груза не особенно высока (r=+0,33), но вполне достоверна (n=170; σr=0,0515; t=6,4; P<0,01). В то же время с размером муравья связан вес приносимых в гнездо частиц строительного материала (r=+0,338; n=66; Р<0,01) и живых насекомых (r=+0,412; n=56; Р<0,01), но не связан вес старых трупов, подобранных на кормовом участке (r=-0,127; n=54; Р>0,05). Это вполне естественно, так как строительный материал (хвоя, веточки) всегда имеется в избытке и, следовательно, размер груза определяется главным образом предпочтением особи. Максимальный размер живой добычи зависит от силы охотника, т.е. в конечном счете также от его размеров. Трупы насекомых, составляющие значительную часть рациона этих муравьев, встречаются на территории достаточно редко и поэтому любой из них, будучи обнаруженным, сразу же приносится в гнездо.
Formica rufa было собрано 170 муравьев, несущих грузы. У каждого муравья измерялась длина груди, а груз сразу же взвешивался на торзионных весах. Результаты измерений показаны на графиках рис:36. В целом корреляция между размером насекомого и весом груза не особенно высока (r=+0,33), но вполне достоверна (n=170; σr=0,0515; t=6,4; P<0,01). В то же время с размером муравья связан вес приносимых в гнездо частиц строительного материала (r=+0,338; n=66; Р<0,01) и живых насекомых (r=+0,412; n=56; Р<0,01), но не связан вес старых трупов, подобранных на кормовом участке (r=-0,127; n=54; Р>0,05). Это вполне естественно, так как строительный материал (хвоя, веточки) всегда имеется в избытке и, следовательно, размер груза определяется главным образом предпочтением особи. Максимальный размер живой добычи зависит от силы охотника, т.е. в конечном счете также от его размеров. Трупы насекомых, составляющие значительную часть рациона этих муравьев, встречаются на территории достаточно редко и поэтому любой из них, будучи обнаруженным, сразу же приносится в гнездо.
Из сказанного ясно, что увеличение вариабельности размеров особей в пределах одной семьи неизбежно должно повлечь за собой расширение спектра добычи и, следовательно, увеличение потенциальной продуктивности кормового участка. Особенно сильно это должно отразиться на видах, значительную долю в питании которых составляют семена (возможность свободного выбора размера груза) и живые насекомые, и в меньшей степени — на видах, питающихся главным образом трупами беспозвоночных. В условиях, когда корма достаточно, это преимущество несущественно, однако в условиях недостатка пищи может оказаться одним из основных способов повышения интенсивности использования кормового участка. Поэтому неслучайно, что высокая вариабельность рабочей касты так часто наблюдается у видов аридных областей. При этом наиболее изменчивыми оказываются крупные карпофаги (Messor aralocaspius, IV=1,92; M. intermedius — 1,82; M. denticulatus и M. variabilis — 1,80), ночные зоофаги (Camponotus xerxes, IV=1,78; C. turcestanus — 1,52; C. turcestanicus — 1,93; C. fedtschenkoi — 1,74) и наиболее крупные зоонекрофаги (Cataglyphis setipes, IV=2,04; C. pallida — 2,0; C. foreli — 1,90; ![]() C. aenescens — 1,86) тогда как у некоторых мелких карпофагов (Tetramorium, Monomorium barbatulum, M. kusnezovi, IV=l,l—1,25) и мелких зоонекрофагов (Plagiolepis, Acantholepis, Tapinoma, IV=1,1—1,2; Cataglyphis emeryi — 1,30) рабочие довольно мономорфны. В гумидной зоне высокая вариабельность рабочей касты наблюдается у видов, численность семей которых превышает несколько десятков тысяч (
C. aenescens — 1,86) тогда как у некоторых мелких карпофагов (Tetramorium, Monomorium barbatulum, M. kusnezovi, IV=l,l—1,25) и мелких зоонекрофагов (Plagiolepis, Acantholepis, Tapinoma, IV=1,1—1,2; Cataglyphis emeryi — 1,30) рабочие довольно мономорфны. В гумидной зоне высокая вариабельность рабочей касты наблюдается у видов, численность семей которых превышает несколько десятков тысяч (![]() Lasius flavus, IV=l,69; Formica s. str. — 1,55—2,04), что несомненно сопровождается необходимостью значительного повышения интенсивности использования кормового участка. Единственное исключение — виды рода Camponotus гумидных областей (C. herculeanus, IV=1,70). Однако поскольку в настоящее время этот род очень широко распространен по всему земному шару и уже в эоцене (балтийский янтарь) рабочие этих муравьев были изменчивыми, невозможно ничего сказать о том, в каких условиях шло становление этого рода.
Lasius flavus, IV=l,69; Formica s. str. — 1,55—2,04), что несомненно сопровождается необходимостью значительного повышения интенсивности использования кормового участка. Единственное исключение — виды рода Camponotus гумидных областей (C. herculeanus, IV=1,70). Однако поскольку в настоящее время этот род очень широко распространен по всему земному шару и уже в эоцене (балтийский янтарь) рабочие этих муравьев были изменчивыми, невозможно ничего сказать о том, в каких условиях шло становление этого рода.
Известно значение разнообразия размеров особей в одновозрастных популяциях рыб (правда, возникающего на генетической основе) для увеличения эффективности использования кормовых ресурсов водоема (Поляков, 1960). Интересно, что спектр питания в разнокачественных популяциях рыб расширяется в условиях слабой обеспеченности пищей, а в водоемах, богатых кормами, питание всех особей сходно (Никольский, 1965).
Первая схема эволюции полиморфизма рабочей касты муравьев была предложена К. Эмери (Emery, 1894). По его представлениям, основными этапами этой эволюции были:
1) рабочие мономорфны и не отличаются размерами от самок;
2) увеличение вариабельности рабочих;
3) исчезновение рабочих средних размеров и четкий диморфизм;
4) в результате исчезновения одной из каст возможно возникновение вторичного мономорфизма.
В общем этой схемы придерживалось и большинство других авторов. Э. Вильсон (Wilson, 1953), обнаружив, что биомодальность распределения рабочих в семье всегда сопровождается аллометрией, и изучив многие промежуточные случаи, предложил несколько иную схему эволюции (пунктирными стрелками показаны редкие случаи переходов):
[ris1.gif]
Как уже отмечалось выше, все существующие построения отражают только феноменологическую сторону эволюции и не рассматривают вопроса о начальных причинах возникновения вариабельности рабочей касты и смены типов изменчивости. Это высказывание, очевидно, требует разъяснения. Дело в том, что все авторы связывают полиморфизм рабочей касты с разделением функций между разными кастами. Однако такое рассмотрение вопроса является по сути своей телеологичным, так как для того, чтобы появилось разделение функций, должны появиться касты или достаточно высокая вариабельность рабочих. При таком подходе становится совершенно неясным, почему полиморфизм рабочих не возникает у общественных насекомых, рабочие которых имеют крылья (пчелы, осы), хотя у них отмечены некоторые различия в предпочтение работ у мелких и крупных особей (Wilson, 1971).
Начальные этапы эволюции полиморфизма рабочей касты у муравьев представляются мне следующим образом: исходным типом изменчивости является не строгий мономорфизм со слабой вариабельностью рабочей касты, а нефиксированный мономорфизм, встречающийся у некоторых видов с примитивной социальной организацией. При таком типе изменчивости размеры рабочих в одной семье могут сильно варьировать, что определяется не какими-то особыми механизмами, а просто зависит от различий в обеспеченности кормом в течение сезона. Еще В. M. Вилер (Wheeler, 1933) отмечал, что у австралийских Myrmecia, наиболее примитивных из ныне живущих муравьев, размер рабочих связан с различиями в обилии пищи в разные сезоны. Позднее это было показано более детальными исследованиями Б. Грея (Gray, 1971а), показавшим различия размеров рабочих Myrmecia dispar зимой и летом и также связавшим эти различия с обеспеченностью семьи кормом. Такая же закономерность обнаружена и у примитивных общественных пчел-галиктин из рода Lasioglossus. При этом оказалось, что наибольшая сезонная изменчивость (5,08%) наблюдается у наиболее примитивно организованных видов (L. zephirum), а по мере усложнения социальности сезонная вариабельность рабочих уменьшается (у L. imitatus — 0,60%) (Breed, 1975). Такая изменчивость хорошо известна и у одиночных насекомых. Различия в пропорциях между мелкими и крупными особями могут быть, но могут и отсутствовать, причем первое, как уже говорилось выше, — отнюдь не свидетельство в пользу появления модификационного полиморфизма.
По мере развития социальности развиваются и совершенствуются механизмы, обеспечивающие оптимальное развитие всех особей, достигающих имагинального состояния, вне зависимости от колебаний условий среды. Возникает строгий, или фиксированный, мономорфизм, при котором все рабочие имеют приблизительно одинаковые размеры.
У большинства видов муравьев средней полосы европейской части СССР, у большинства социальных пчел и ос наблюдается именно такой тип изменчивости. Промежуточным будет состояние, когда все развивающиеся личинки получают одинаковое количество корма, но в зависимости от обеспеченности пищей средний размер рабочих в разных семьях будет различаться, что свидетельствует о несовершенстве социального гомеостаза. Подобные различия между семьями обнаружены у ![]() Myrmica rubra (Brian, Brian, 1951) и ряда других видов.
Myrmica rubra (Brian, Brian, 1951) и ряда других видов.
В пользу того, что фиксированный мономорфизм определяется не генетическими причинами, а социальной регуляцией, свидетельствует тот факт, что у всех мономорфных видов, самки которых самостоятельно основывают новые семьи, первые рабочие (микрэргаты, по терминологии Вилера) значительно меньше нормальных. Одним из демпферных механизмов, обеспечивающих стабильность выкармливания расплода, является каннибализм, широко распространенный у всех социальных перепончатокрылых (обзор: Flanders, 1970). У Myrmica ussuriensis, например, не менее 20—50% яиц и молодых личинок скармливается рабочими развивающимся личинкам (Dlussky, Kupianskaya, 1972). Другой хорошо известный механизм — создание запасов пищи для расплода (семян, пыльцы, высушенных насекомых).
Фиксированный мономорфизм является устойчивым состоянием, достигнутым в результате длительной эволюции социальности. Повышение вариабельности должно быть связано с нарушением хорошо отработанной системы поддержания социального гомеостаза, что в обычных условиях привело бы к снижению адаптивности. Небольшие различия в предпочтении выбора работ у особей разного размера, как показывает пример высших пчел, не могут нарушить этого состояния и привести к появлению полиморфизма. Однако в условиях недостатка корма может возникнуть отбор на разнообразие рабочей касты, приводящий к перебалансировке системы социального гомеостаза и появлению фиксированной вариабельности, при которой рабочие одной семьи всегда сильно отличаются размерами. Несомненно, что различия в размерах определяются разными режимами выкармливания личинок. Можно предположить, что это могут быть различия в количестве пищи, различия в количестве скармливаемых секретов желез или ингибиторы роста, добавляемые в пищу части личинок. Условия, благоприятные для развития повышенной вариабельности рабочей касты, могут возникнуть при общей бедности биогеоценоза (например, в пустынях), при высокой интенсивности конкуренции и ограниченности кормовой базы вследствие высокой ярусной специализации (например, в тропических лесах) или при значительном повышении численности семьи и ограниченности кормовых ресурсов на кормовом участке (многие виды с высоко развитой социальностью).
При таком подходе становится ясным, почему полиморфизм рабочей касты не мог возникнуть у пчел: увеличение вариабельности в размерах у насекомых, питающихся нектаром и пыльцой, практически не может отразиться на расширении спектра питания. У складчатокрылых ос размеры фуражира также не связаны с размерами добычи, так как, во-первых, даже небольшая оса может убить довольно крупное насекомое, а, во-вторых, и это, по-видимому, основное, добыча расчленяется на месте и в гнездо доставляются не трупы целиком, а комки пережеванных тканей.
Некоторые затруднения для предложенной гипотезы представляет тот факт, что у мелких карпофагов пустынь Средней Азии и Северной Африки рабочие мономорфны (у Tetramorium inerme, Monomorium barbatulum и M. kusnezovi, IV=1,1; У T. schneideri — 1,25). Кажется, мономорфизм рабочих мелких карпофагов может быть объяснен только в том случае, если рассматривать эти виды как элемент коадаптивного комплекса. В большинстве пустынь Палеарктики доминирующей группой карпофагов являются муравьи-жнецы из рода Messor. Рабочие этих муравьев весьма вариабельны, и самые мелкие особи ненамного крупнее мелких карпофагов: длина груди рабочих Tetramorium schneideri — 0,9—1,25 мм, различных видов Monomorium — 0,6—1,0 мм, a Messor aralocaspius — 1,3—2,5 мм. Таким образом, на долю мелких карпофагов остаются лишь самые мелкие семена. Уменьшение размеров фуражиров при этом нецелесообразно, так как не расширяет спектра питания этих муравьев, а увеличение повлекло бы за собой усиление конкуренции с муравьями-жнецами.
Эту точку зрения подтверждает то, что в пустынях Индостана, где Messor (группа M. hymalayanus), видимо, появились сравнительно недавно и не являются доминирующей группой, широко распространены мелкие карпофаги с вариабельными рабочими (Monomorium, Pheidole). Среди пустынных мелких карпофагов Средней Азии и Северной Африки вариабельность рабочих отмечается только у Pheidole pallidula (диморфизм) и ![]() Monomorium gracillimum (IV=1,74). Оба эти вида несомненно появились здесь недавно и скорее всего происходят из пустынь или саванн Юго-Восточной Азии.
Monomorium gracillimum (IV=1,74). Оба эти вида несомненно появились здесь недавно и скорее всего происходят из пустынь или саванн Юго-Восточной Азии.
В пустынях Северной Америки мелкие карпофаги представлены главным образом видами рода Pheidole с вариабельными или даже диморфными рабочими, причем в отсутствие муравьев-жнецов Pogonomyrmex численность Pheidole сильно возрастает (Schumacher, Whitford, 1976). Довольно вариабельны и рабочие Tetramorium пустынь южной Африки (Stitz, 1923), где отсутствуют муравьи-жнецы.
Фиксированная вариабельность с унимодальным частотным распределением особей по размерным классам и монофазной аллометрией является эволюционно неустойчивым состоянием. Прежде всего при ее появлении выявляются различия в предпочтении работ мелких и крупных особей. Это хорошо проиллюстрировал Т. Шнейрла (Schneirla, 1971) на примере Eciton: мелким рабочим гораздо удобнее ухаживать за яйцами, тогда как переноску куколок во время рейдов гораздо легче осуществляют крупные особи. Эти различия могут послужить основой для дальнейшей функциональной дифференциации. При этом у мелких рабочих могут оказаться выгодными одни признаки, а у крупных — другие. Как это обычно происходит в аналогичных случаях с адаптивными модификациями (Шмальгаузен, 1969), возникают разные каналы онтогенеза, что будет отражаться на характере аллометрии (она становится полифазной). Включение одного из двух (или более) каналов онтогенеза обычно происходит при помощи какого-либо простого сигнала (у общественных насекомых — трофического или гормонального). Неизбежным следствием далеко зашедшей канализации онтогенезов будет постепенное исчезновение промежуточных форм и, следовательно, появление полиморфизма.
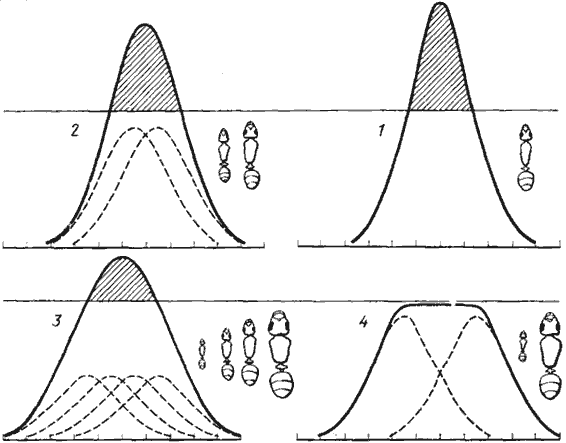
Второй причиной возникновения полиморфизма является то, что при низкой обеспеченности кормом особи средних размеров начинают работать в неоптимальном режиме. Поясню это на упрощенной схеме (рис:37). Представим себе, что фуражиры каждого размерного класса работают в оптимальном режиме и собирают добычу восьми размерных классов, причем частотное распределение количества единиц добычи разных размерных классов подчиняется биномиальному распределению (соотношение 1:7:21:35:35:21:7:1). Таким же, естественно, будет распределение добычи по размерам в семье с мономорфными рабочими (рис:37/1/). При некотором увеличении вариабельности (появление двух близких размерных классов фуражиров) и сохранении того же общего количества единиц добычи суммарная кривая становится несколько шире и соответственно менее эксцессивной (рис:37/2/). Еще ниже и шире будет суммарная кривая при четырех размерных классах фуражиров (рис:37/3/). Если мы теперь представим себе, что количество единиц пищи каждого размерного класса на кормовом участке ограничено (горизонтальная линия на всех схемах), окажется, что для того чтобы получить то же количество единиц, муравьи будут собирать часть добычи более мелкого или более крупного размера и соответственно работать в неоптимальном режиме. Этого можно избежать, ликвидировав рабочих средних размеров при сохранении той же общей численности. При этом эксцессивность суммарной кривой частотного распределения добычи по размерным классам резко падает, а при определенных условиях вершина ее становится плоской (рис:37/4/). При этом, однако, широта спектра добычи не изменяется.
В предложенных упрощенных моделях численность фуражиров разных размерных классов принималась одинаковой, но в естественных условиях этого, конечно, не наблюдается. На начальных этапах эволюции вариабельности численность рабочих средних размеров будет выше, чем мелких или крупных, что еще больше увеличивает эксцессивность суммарной кривой. Для упрощения также было принято, что единицы добычи разных размерных классов равноценны, но на самом деле, конечно, это не так. Поскольку, кроме того, в естественных условиях крупной добычи всегда меньше, чем мелкой, численность крупных рабочих должна быть меньше, чем мелких. Именно такая картина и наблюдается у большинства полиморфных видов.
Подводя итоги, хотелось бы отметить, что эволюцию полиморфизма рабочей касты у муравьев можно рассматривать как один из примеров квантовой эволюции, правда, несколько отличающийся от классических случаев, рассматриваемых Дж. Симпсоном (1948). Имеются два устойчивых состояния — мономорфизм и диморфизм. Определенные внешние условия (недостаток пищи на кормовом участке) могут нарушить устойчивость мономорфизма и спровоцировать повышение вариабельности. Однако такое состояние является неустойчивым и довольно быстро фиксированная вариабельность может перейти в диморфизм. Очевидно, переходными состояниями являются слабая двухфазная аллометрия при сохранении унимодальности частотного распределения (некоторые Messor) и нечеткая бимодальность при сохранении монофазной аллометрии (некоторые Cataglyphis). Обратный переход от диморфизма к мономорфизму может осуществиться только за счет исчезновения одной из форм. Как в процессе эволюции происходит изменение соотношения каст и исчезновение одной из них, объясняют модели Э. Вильсона (Wilson, 1968, 1971), но обсуждение их выходит за рамки данной работы.
Хотя увеличение вариабельности рабочей касты мы замечаем по морфологическим признакам, вряд ли эта адаптация может рассматриваться как морфологическая. Как уже отмечалось выше, онтогенетические потенции к повышенной вариабельности имеются у многих мономорфных видов, на что указывает наличие микрэргатов. В то же время переход от фиксированного мономорфизма к фиксированной вариабельности осуществляется за счет изменения поведения муравьев при уходе за потомством. Переход от фиксированной вариабельности к полиморфизму сопровождается изменением характера онтогенеза различных подкаст и может рассматриваться как морфофизиологическая адаптация.
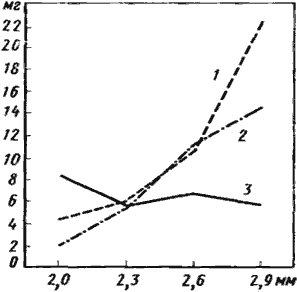
Рис. 36. Корреляция между размерами рабочих особей
1 — гнездовой материал; 2 — свежеубитые беспозвоночные; 3 — трупы беспозвоночных
в биологии (от др.-греч. πολύμορφος - многообразный) - способность некоторых организмов существовать в состояниях с различной внутренней структурой или в разных внешних формах.
У муравьев это проявляется в том, что представители разных каст морфологически очень сильно отличаются друг от друга. Отсюда условное деление на «рабочих» и «солдат».
необратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.
(rp. ontos - сущее, существо) - индивидуальное развитие организма с момента зарождения до смерти.
у муравьев имеется три основные касты: самцы, самки и рабочие (бесплодные модифицированные самки).
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
живут колониями (пчелы, муравьи, термиты, некоторые осы). В состав колонии, напр. у термитов, входят половые особи (самцы и матки обеспечивают размножение), солдаты (защиту) и рабочие (собирание корма, строительство гнезда и т. д.). Солдаты и рабочие бесплодны; половые особи и солдаты сами не способны питаться; таким образом, члены колонии полностью зависят друг от друга. Разделение функций и сложные взаимодействия разных особей в колониях общественных насекомых обеспечиваются рядом особенностей в их строении и поведении; последнее регулируется с помощью выделяемых ими химических веществ (феромонов).
функциональное образование, состоящее из материнской и дочерних семей, поддерживающих регулярные родственные отношения
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
Старое название: Pheidole arenarum Ruzsky
Всего 106 видов.
 Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравейучасток, используемый муравьями для добычи пищи.
животные, не имеющие позвоночника: простейшие, губки, кишечнополостные, черви, моллюски, членистоногие, иглокожие и др.; примерно 3/4 видов всех животных.
организм, питающийся семенами растений.
ксеробионт
гигрофил
организм, питающийся исключительно другими животными
(зоонекрофаг), организм, питающийся мертвыми животными, гетеротроф
ксеробионт
ксеробионт
 Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунок
Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунокгигрофил, галобионт
Старое название: Cataglyphis flavigastra
Всего 415 видов
галобионт
Subspecies: Monomorium indicum kusnezowi Santschi, 1928
Всего 60 видов.
Всего 62 вида.
ксеробионт
Старое название: Proformica deserta
 Lasius flavus (Fabricius, 1782) — желтый луговой (садовый, земляной) муравей
Lasius flavus (Fabricius, 1782) — желтый луговой (садовый, земляной) муравейВсего 965 видов.
совокупность особей одного вида, длительно занимающая определенное пространство и воспроизводящая себя в течение большого числа поколений. В современной биологии популяция рассматривается как элементарная единица процесса эволюции, способная реагировать на изменения среды перестройкой своего генофонда.
надсемейство летающих насекомых отряда перепончатокрылых, родственное осам и муравьям. Существует около 20 тысяч видов пчёл. Распространены на всех континентах, кроме Антарктиды. Пчёлы питаются нектаром и пыльцой, используя нектар главным образом в качестве источника энергии, а пыльцу для получения белков и других питательных веществ.
сборное название насекомых подотряда жалящих перепончатокрылых (кроме пчел и муравьев). Длина до 40 мм (шершни). Ок. 23 тыс. видов. Распространены широко. Опылители растений. Личинки некоторых ос паразиты других насекомых.
Всего 90 видов.
обязательное существование только сообществами (семьями, общинами).
 Myrmica rubra (Linnaeus, 1758) — рыжая мирмика
Myrmica rubra (Linnaeus, 1758) — рыжая мирмикаSubspecies: Myrmica scabrinodis ussuriensis, Kuznetsov-Ugamsky
разноцветные
 (белые, желтые,оранжевые, коричневые) комочки неправильной формы, ~2-3мм диаметром, обножка это то что пчелы приносят на лапках. Слегка сладковатая на вкус с ярким цветочным привкусом. Пыльца — природный концентрат, содержащий белки, все известные витамины, ценные минеральные вещества, по общему аминокислотному составу обножка близка к другим богатым белком пищевым продуктам — мясу, молоку, яйцам.
(белые, желтые,оранжевые, коричневые) комочки неправильной формы, ~2-3мм диаметром, обножка это то что пчелы приносят на лапках. Слегка сладковатая на вкус с ярким цветочным привкусом. Пыльца — природный концентрат, содержащий белки, все известные витамины, ценные минеральные вещества, по общему аминокислотному составу обножка близка к другим богатым белком пищевым продуктам — мясу, молоку, яйцам.эволюционно сложившаяся, пространственно ограниченная, длительно самоподдерживающаяся, однородная экологическая система, в которой функционально взаимосвязаны живые организмы и окружающая их абиотическая среда.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
Всего насчитывает 549 видов.
 Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушитель
Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушительСтарое название: Monomorium gracillimum
Всего 60 видов.
Всего 12 видов.
Всего 65 видов.






