






В пустынях Средней Азии встречаются две группы видов рода Messor, сильно отличающиеся экологически. Представители первой группы, куда относятся M. variabilis, M. excursionis и M. lamellicornis, собирают пищу исключительно ночью, практически не организуют групповой фуражировки и собирают преимущественно одиночные рассеянные семена. Представители второй группы, куда относится подавляющее большинство видов рода, работают преимущественно в светлое время суток (исключение — M. intermedius) и, Как правило, фуражируют большими группами на полях эфемеров или однолетних солянок. Обычно в каждом биотопе встречается один вид из первой группы и один-два вида — из второй. Сходная картина наблюдается и в пустынях Северной Америки (Davidson, 1977а).
У M. variabilis все фуражиры являются одиночными сборщиками. Обычно выход муравьев начинается с наступлением темноты и к утру заканчивается. Максимальное расстояние от гнезда, на котором мы обнаруживали фуражиров этого вида, — 20 м, но вполне возможно, что они могут уходить и на большее расстояние (по косвенным данным — не менее 50 м). Динамическая плотность фуражиров на территории очень низка — менее 0,01 особей/дм²/мин. Обычно при выставлении кормушки на участке M. variabilis в нескольких метрах от гнезд за ночь ее обнаруживали один-два рабочих, которые и работали до утра. В одном опыте кормушка была оставлена на несколько дней. 23 августа 1971 г. в Акыбае в 4 м от гнезда этого вида было выложено несколько кучек риса. До 2 ч ночи муравьи приманки не обнаружили. 24 августа наблюдения не проводились, но к утру 25 августа приманка еще была на месте. В 20 ч 30 мин муравьи вышли из гнезда и сразу несколько рабочих направились к приманке. Всего в эту ночь работало семь особей, причем все они ходили поодиночке с большими интервалами.
Совсем иная картина наблюдается у M. aralocaspius, M. denticulatus, M. intermedius и многих других видов рода, обитающих в пустынях Средней Азии. Как правило, фуражировка происходит большими группами особей. От гнезда отходят одна или две дороги, по которым муравьи выходят на участки созревших растений. Длина дорог может достигать 100 м, но обычно она бывает порядка 10 м. На кормовом участке муравьи либо собирают семена на земле, либо откусывают их, поднявшись на растения. К гнезду почти все фуражиры идут с семенами (рис:82). Если на разных дорогах прометить муравьев, то вскоре обнаруживается, что они не привязаны к какой-либо территории и свободно переходят с дороги на дорогу. Обычно муравей выходит из гнезда на площадку около входа и дожидается прихода муравьев с семенами, а затем идет в том направлении, откуда пришла первая группа. Контактов между ними при этом обычно не наблюдается. По пути он ориентируется по потоку фуражиров, идущих с кормового участка.
Каждый раз при выходе муравьев после ночного или дневного перерыва фуражировки дороги формируются заново. В идеальном случае, т.е. при отсутствии конкурирующих семей и при равномерном распределении пищи на территории, муравьи последовательно собирают урожай созревших семян в разных секторах кормового участка, так что за один цикл делается полный круг (рис:83). Длительность цикла колеблется у M. aralocaspius в период созревания семян в пределах одной — трех недель. При этом циклы разных семей не совпадают, так что обычно одновременно работает только часть гнезд. По окончании цикла муравьи закрывают гнездо и несколько дней не появляются на поверхности. Пример такого цикла подробно описан в работе А. А. Захарова (1972). В других случаях правильность смены секторов может нарушаться. Но всегда для M. aralocaspius, M. denticulatus и M. intermedius в пустынях характерно чередование периодов фуражировки и периодов покоя.
Переход из одного состояния в другое определяется главным образом двумя факторами — количеством пищи на кормовом участке и размерами запасов в гнезде. Так, например, в августе — сентябре 1971 г. на опытной площадке в Акыбае созревших семян практически не было и все гнезда M. aralocaspius были закрыты. В конце августа мы выложили здесь несколько приманок из зерна. Это вызвало выход муравьев из части гнезд и организацию групповой фуражировки. 29 апреля 1970 г. около двух гнезд M. denticulatus были выставлены кормушки с пшеном. На следующий день интенсивность фуражировки в этих гнездах возросла в несколько раз, а через 4 и 7 дней муравьи заполнили все камеры и прекратили фуражировку. В это же время в соседних гнездах, не получивших подкормки, фуражировка не прекратилась (рис:48).
В пустыне муравьи-жнецы используют единовременно не более 20% от своего потенциального кормового участка (Сапарлиев, 1973). При этом участки, на которых работают муравьи из разных семей, никогда не перекрываются. В то же время полные кормовые участки соседних гнезд, определенные последовательным картированием за длительное время, могут перекрываться (рис:84). Таким образом можно сказать, что у этих муравьев территории перекрываются в пространстве, но изолированы во времени. Столкновения между фуражирами из разных гнезд наблюдаются очень редко. В том случае, когда они все же происходят и фуражиры разных семей одного вида выходят на одно и то же поле, у M. aralocaspius и M. denticulatus настоящих схваток не бывает. Муравьи становятся друг против друга в угрожающей позе, открыв челюсти и высоко приподнявшись на ногах (рис:82). При этом они мелко дергаются всем телом и время от времени делают выпады (скорее, прыжки) в направлении друг друга. Такой ритуал продолжается 10—15 мин, после чего муравьи расходятся и между кормовыми участками устанавливается нейтральная зона.
При столкновении разных видов муравьи ведут себя более агрессивно. В мае 1978 г. мы наблюдали такое столкновение между M. aralocaspius и M. rufitarsis darianus в Варганзе. На территории нашего лагеря рядом с тем местом, где мы постоянно обедали, находилось гнездо M. rufitarsis. Резкое увеличение количества корма вокруг этого гнезда (рассыпанная крупа, крошки) привело к тому, что от находящегося в 7 м гнезда M. aralocaspius образовалась тропа в этот участок. Муравьи блокировали гнездо M. rufitarsis. На гнездовом валу этого гнезда постоянно находилось около 50 рабочих M. aralocaspius, главным образом крупных. Во входе в гнездо M. rufitarsis собралась группа крупных рабочих, которые отгоняли пришельцев (рис:85). Время от времени один из них нападал на пришельцев. При этом муравьи сцеплялись мандибулами и так часами стояли или медленно тянули друг друга. Остальные муравьи лишь изредка подходили к такой паре, принимали угрожающую позу, ощупывали их антеннами и отходили в сторону. Это могло продолжаться несколько часов (одну пару наблюдали 4 ч), после чего муравьи расходились. Несколько рабочих M. aralocaspius пытались проникнуть в поверхностные камеры M. rufitarsis, сделав подкопы в 20—30 см от входа. Когда один из подкопов (всего их было три, но два муравьи не закончили) достиг камер гнезда, там завязалась короткая схватка, в результате которой M. aralocaspius отступили и прекратили рыть почву. Цикл суточной активности этих видов несколько не совпадает. К 18 ч M. aralocaspius обычно уходили и M. rufitarsis выходили из гнезда и собирали корм в радиусе не более 1 м. Такое положение продолжалось три дня, после чего M. aralocaspius ушли и восстановились прежние нейтральные отношения.
Организацию групповой фуражировки у муравьев-жнецов осуществляют активные фуражиры, которые составляют всего 3% от общего числа фуражиров. Их поведение резко отличается от поведения пассивных фуражиров. Как правило, они работают как одиночные сборщики и не участвуют в коллективной работе на полях эфемеров. Все они имеют индивидуальные поисковые участки площадью 0,2—0,4 м². Индивидуальные участки разных особей могут перекрываться (рис:86). Активные фуражиры постоянно обследуют весь потенциальный кормовой участок семьи и за счет этого осуществляется постоянный контроль за ходом созревания семян.
В типичном случае образование дороги происходит у M. aralocaspius, M. denticulatus и M. intermedius сходным образом. Когда условия на кормовом участке становятся благоприятными для фуражировки (с наступлением рассвета или со снижением температуры почвы до 35—39° у первых двух видов или с наступлением сумерек у M. intermedius), из гнезда появляется большое число муравьев. Все они скапливаются вблизи входа и держатся в пределах гнездового вала, т.е. в круге радиусом 30—60 см. Через 10—15 мин после начала массового выхода в круге скапливается 150—300 особей (у M. intermedius — больше). В какой-то момент из этой массы выбегает несколько муравьев, среди которых находится один или несколько активных фуражиров. Пробежав около 50 см, эти муравьи обычно возвращаются. Но за ними уже устремляются пассивные фуражиры. Поведение этих муравьев в общем сходно. Каждый из них стремится уйти от гнезда в том направлении, откуда возвращаются другие муравьи. Дойдя до того места, где поток кончается, он пробегает еще несколько сантиметров и, обнаружив, что навстречу и впереди никто не идет, быстро поворачивает обратно. Но он уже сам, возвращаясь, служит ориентиром для идущих за ним особей, которые проходят немного дальше и также поворачивают назад. Оказавшись среди гущи муравьев, пассивный фуражир снова поворачивается и бежит от гнезда до тех пор, пока снова не окажется за пределами колонны. В результате этих челночных пробежек отдельных особей весь поток медленно продвигается вперед в направлении, заданном активными фуражирами. Скорость его движения обычно 15— 20 см/мин, тогда как скорость отдельных рабочих достигает 1—3 м/ /мин.
Изъятие отдельных муравьев, идущих первыми, не приводит к остановке общего движения. В одном из опытов (28.IV.70, гнездо МД-10, M. denticulatus) мы изъяли из головы потока на расстоянии 75 см от гнезда 50 первых особей, но, несмотря на это, движение продолжалось. В другом опыте (27.IV.70, гнездо МА-12, M. aralocaspius) мы смели в сторону часть муравьев вместе с верхним слоем почвы, но через минуту продвижение колонны восстановилось в прежнем направлении. Однако если на пути потока возникает непреодолимое препятствие, дорога не образуется. Так, 26 апреля 1970 г. мы в 16 ч затенили гнездо МА-21 M. aralocaspius и прилежащий участок такыра. В 17 ч (на час раньше, чем в других гнездах) сформировался поток, который вскоре достиг границы света и тени. Температура на поверхности почвы на солнце была еще выше 40°, и все муравьи сразу же поворачивали, достигнув границы. Вскоре движение прекратилось, муравьи ушли в гнездо и больше в этот день не выходили для групповой фуражировки.
Дойдя до участка с созревшими семенами, фуражиры, идущие первыми, быстро находят семена и возвращаются в гнездо. Другие муравьи рассеиваются по участку и начинают сбор. Устанавливается дорога. Первые фуражиры с семенами начинают возвращаться в гнездо обычно уже через 10—15 мин после начала мобилизации. Изредка поток фуражиров изменяет направление движения. Один из таких случаев описан А. А. Захаровым (1972). Пройдя 2—3 м, колонна M. aralocaspius дошла до участка, бедного семенами, и лишь часть муравьев рассеялась для сбора. В голове потока образовалось большое скопление рабочих. Вскоре здесь появились разведчики со свитой, которые отделились от скопления и пошли почти под прямым углом к первоначальному направлению. Значительная часть пассивных фуражиров отправилась за ними. В результате образовалась дорога с резким изгибом, которую муравьи впоследствии спрямили (рис:87). В ряде случаев мы наблюдали одновременное формирование двух дорог.
Поскольку относительная численность активных фуражиров в гнезде очень низка, общая площадь кормового участка велика и заранее предугадать, куда направится дорога, невозможно, нам не удавалось заранее промаркировать активных фуражиров, участвующих в организации групповой фуражировки в естественных условиях и провести детальные наблюдения за их поведением. Это удалось сделать лишь в опытах с выкладыванием приманок. На одном из этих опытов следует остановиться подробнее, так как он одновременно иллюстрирует несколько характерных особенностей поведения активных фуражиров. Утром 29 апреля 1970 г. в Акыбае в 1 м от гнезда M. denticulatus МД-4 была выставлена кормовая площадка без приманки. Все муравьи, проходившие через эту площадку, были промечены индивидуальными метками, а затем были закартированы индивидуальные поисковые участки меченых рабочих (рис:86/вверху/).
В 17 ч 50 мин, когда начался вечерний выход из гнезда, но дороги еще не сформировались, на кормовую площадку было выложено 65 г пшена. В 17 ч 55 мин обычным способом сформировалась дорога А на восток от гнезда (рис:86/Л/). В 17 ч 57 мин приманку обнаружил фуражир 020 и понес зерно в гнездо. В 18 ч корм обнаружил 022, в 18 ч 04 мин — 011 и 200. Все эти муравьи также вернулись в гнездо с обнаруженными семенами. В 18 ч 07 мин из гнезда вышла группа из 30 особей, во главе которой шли разведчики 022 и 011. Следом за ними выходили все новые и новые рабочие. Примерно на половине дороги активные фуражиры отстали от потока и на кормушку первыми вышли немеченые особи. 022, дойдя до кормушки, походил по площадке и, не взяв зерна, направился в свой индивидуальный участок, где и занимался сбором семян эфемеров до конца опыта. 011 сделал еще один рейс с пшеном ив 18 ч 17 мин также ушел в свой участок. Муравей 010 до начала групповой фуражировки занимался сбором семян эфемеров, но когда через его индивидуальный участок пошла дорога, он, захваченный общим потоком, дошел до кормушки, но сразу же ушел на новый участок (010а, рис:86), где и собирал семена эфемеров до конца опыта. Из всех активных фуражиров лишь 200, на индивидуальном кормовом участке которого находилась кормушка, совершил около десяти рейсов с пшеном и последний раз был отмечен здесь в 19 ч 07 мин.
В 18 ч 11 мин большая группа пассивных фуражиров прошла мимо кормушки и сразу же описанным выше способом начала формироваться дорога В на север от гнезда. Пройдя 3 м, муравьи в 18 ч 38 мин обнаружили небольшой довольно бедный участок с эфемерами и некоторые из них понесли в гнездо семена. Но здесь фуражировали рабочие из соседнего гнезда МД-6 и между муравьями начались отдельные стычки. До 18 ч 50 мин на дороге В не было ни одного меченого рабочего. В 18 ч 50 мин появился муравей 011, который организовал поворот дороги и увлек за собой других муравьев. В 18 ч 55 мин дорога подошла почти к самому гнезду МД-6 и повсюду начались ритуальные схватки (рис:82). В 19 ч 08 мин муравьи из гнезда МД-4 повернули назад и к 19 ч 30 мин дорога В прекратила свое существование. К этому же времени закончилось движение и по дороге А, так что муравьи работали только на кормушке.
Таким образом, на примере этого опыта видно, что именно активные фуражиры задают направление потоку пассивных фуражиров при образовании дороги. Причем явственно видно, что они именно задают направление, а не ведут пассивных фуражиров. Такая система мобилизации необычайно эффективна, так как позволяет немногим активным фуражирам вывести на кормовой участок, расположенный за несколько (а иногда — несколько десятков) метров от гнезда, несколько сот пассивных фуражиров за один раз. Поскольку следовые феромоны при мобилизации не используются, она может быть произведена и после значительного перерыва, во время которого почва нагревается до 50— 70°. В то же время такой способ мобилизации не отличается особой точностью наведения. В том случае, когда источник пищи занимает небольшую площадь (кормушка), могут происходить ошибки, что и иллюстрирует описанный опыт. Однако в естественных условиях, когда мобилизация производится на участок территории семьи с созревшими эфемерами, высокая точность наведения не нужна: Интересен и тот факт, что активные фуражиры, проведя мобилизацию, не участвуют в групповой фуражировке, а продолжают работать как одиночные сборщики на своих индивидуальных участках или даже (010) переходят на другой участок в стороне от дороги.
Помимо описанного обычного способа, у муравьев-жнецов изредка наблюдается и мобилизация при помощи пахучего следа. Это происходит только на небольшом расстоянии от гнезда. Так, 30 апреля 1970 г. в Акыбае в 50 см от гнезда M. aralocaspius MA-18 была выложена приманка из пшена. К этому времени муравьи уже сформировали одну дорогу (рис:88). Почти сразу приманку обнаружил один из активных фуражиров, ходящих вне дороги, взял зерно и направился к гнезду. Перед гнездом мы его убрали. Несмотря на это, от гнезда по следу сразу же направилась группа из 20 муравьев. После того как их убрали и подмели участок, тропа не восстановилась. Через 2 мин кормушку обнаружил второй разведчик. Он, однако, вышел не к гнезду, а на дорогу в 30 см от гнезда. По его следу также сразу же направилось к кормушке множество муравьев. Сам разведчик в это время пошел в гнездо. Мобилизовавшиеся фуражиры двигались не точно по следу, а шли параллельно ему полосой шириной 5—7 см. Часть из них дошла до кормушки, а некоторые прошли дальше и, как и в описанном выше опыте, образовали ложную дорогу. Через некоторое время она исчезла, но до конца наблюдений (в течение часа) дорога от кормушки выходила не к гнезду, а на основную дорогу.
Следует отметить, что все описанные особенности использования кормового участка муравьями-жнецами характерны только для бедных кормом пустынных и полупустынных территорий. Поведение этих муравьев отличается высокой пластичностью. В условиях, когда корма много, например в полусаваннах Бадхыза или в Копетдаге, M. denticulatus и M. intermedius имеют постоянную систему дорог, расположение которых остается постоянным в течение многих дней. При этом все дороги используются муравьями одновременно. Пример такой территории M. denticulatus изображен на рис:89.
Система использования кормового участка, сходная с Messor, наблюдается и у муравьев-жнецов пустынь Северной Америки, представленных видами Pogonomyrmex, Veromessor и Novomessor. Здесь так же, как и в пустынях Средней Азии, на одной территории обычно обитают совместно виды, собирающие рассеянные семена и не организующие групповой фуражировки, и виды, обычно фуражирующие с помощью дорог (Davidson, 1977b). Кормовые участки разных семей перекрываются, но при образовании дорог соседние семьи собирают корм в разных участках (Whitford, Ettershank, 1975; Whitford, 1976). Так же, как и в пустынях Средней Азии, у муравьев-жнецов периоды активности чередуются с периодами покоя, причем авторы работ связывают перерывы активности с заполнением камер с запасами (Whitford, Ettershank, 1975). У Veromessor pergandei, в пустынях Калифорнии довольно регулярно делающего дороги, муравьи ежедневно используют для фуражировки только часть кормового участка. Направление выхода колонны каждый день меняется, причем оно поворачивается постепенно либо по часовой стрелке, либо против (обычно направление поворота характерно для каждого гнезда). Угол поворота тем больше, чем меньше пищи имеется на территории. В период массового созревания семян эти муравьи фуражируют одновременно во всех направлениях от гнезда. В сухой сезон разбрасывание семян по территории V. pergandei вызывало переход к сбору пищи во всех направлениях уже через 2—3 ч (Davidson, 1977b). По имеющимся данным (Holldobler, Wilson, 1971), для организации групповой фуражировки Pogonomyrmex используют пахучий след, но эти данные были получены в лабораторных условиях. Однако следует учитывать, что эксперименты на малых аренах (для муравьев-жнецов — с радиусом менее 2—3 м) воспроизводят только поведение муравьев в пригнездовой зоне. Экспериментов в полевых условиях с американскими муравьями-жнецами пока не проводилось.

Рис. 82. Фуражировка Messor aralocaspius
Внизу слева два муравья в угрожающих позах, которые они обычно принимают при встрече на кормовом участке особей из другой семьи. Внизу справа — рабочий Cataglyphis setipes, основной собиратель трупов M. aralocaspius, вверху справа — жужелица Anthia mannerheimi (изображен самец, мелкий экземпляр, крупные в 1,5 раза больше), основной истребитель муравьев-жнецов в песчаных пустынях Каракумов, в характерной позе при ловле муравьев на фуражировочной дороге.
Рисунок сделан на основании зарисовок и фотографий автора в Акыбае
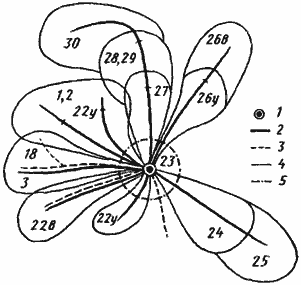
Рис. 83. Использование кормового участка фуражирами гнезда АК-3 Messor aralocaspius в Акыбае в период с 16 апреля по 7 мая 1970 г.
[Захаров, 1972]
1 — гнездо; 2 — дороги с 22 апреля по 3 мая; 3 — дороги до 16 апреля; 4 — границы сбора семян; 5 — зона сбора семян вокруг гнезда. Цифрами обозначены даты фуражировки по секторам, индексами — утренняя (у) и вечерняя (в) фуражировки
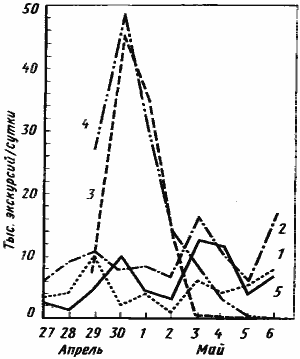
Рис. 48. Влияние избытка пищи на динамику активности Messor denticulatus
(Акыбай, 1970 г.)
1 —5 — номера гнезд (соответственно МД-7001—МД-7005). Рядом с гнездами №3 и 4, 29 апреля были выставлены кормушки с пшеном. Гнезда №1, 2 и 5 служили контролем
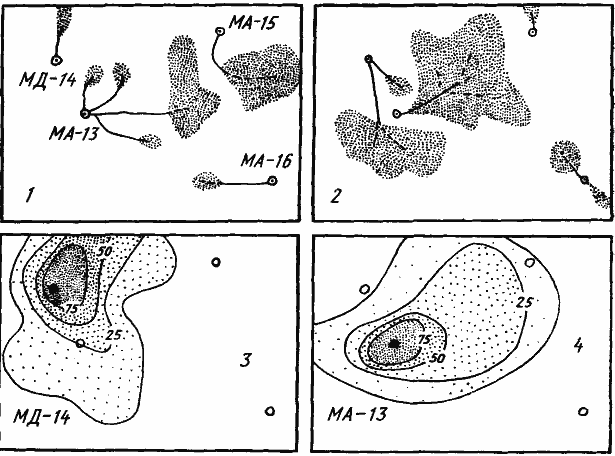
Рис. 84. Кормовые участки разных семей M. aralocaspius (МА-13) и M. denticulatus (МД-14, МД-15, МД-16) на пробной площадке в Акыбае
1 — кормовые участки гнезд 4 апреля; 2 — то же, 25 апреля 1970 г.; 3, 4 — частота посещения (в %) различных участков территории фуражирами семьи МД-14 (3) и семьи МА-13 (4) в течение апреля 1970 г.
Размер площадки 8X11 м

Рис. 85. Нападение рабочих Messor aralocaspius на гнездо M. rufitarsis darianus
Рисунок сделан на основании полевых зарисовок и фотографий в Варганзе в мае 1978 г.
Объяснения в тексте
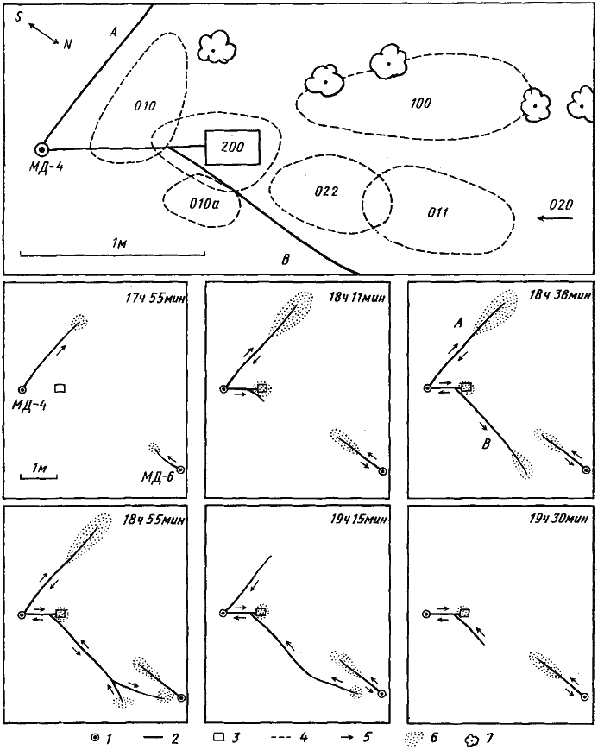
Рис. 86. Индивидуальные поисковые участки активных фуражиров семьи МД-4 Messor denticulatus (верхний рисунок) и последовательные этапы формирования дороги к кормушке с пшеном
(Акыбай, 29 апреля 1970 г.)
1 — гнезда;
2 — дороги;
3 — кормушка с пшеном;
4 — границы индивидуальных поисковых участков активных фуражиров (цифры — код индивидуальных меток);
5 — направления движения пассивных фуражиров по дорогам;
6 — зона рассеяния фуражиров;
7 — кусты тамарикса
Объяснения в тексте
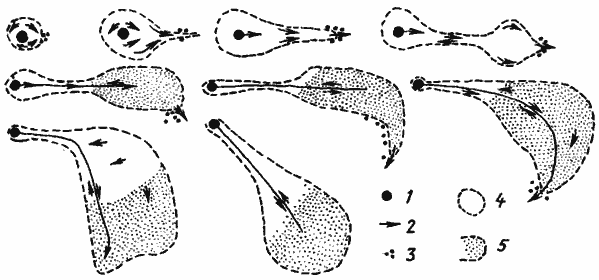
Рис. 87. Последовательные этапы формирования фуражировочной дороги гнезда АК-2 Messor aralocaspius в Акыбае 18 апреля 1970 г.
[Захаров, 1972]
1 — гнездо; 2 — направления движения муравьев; 3 — активные фуражиры; 4 — зона рассева пассивных фуражиров; 5 — зона сбора семян
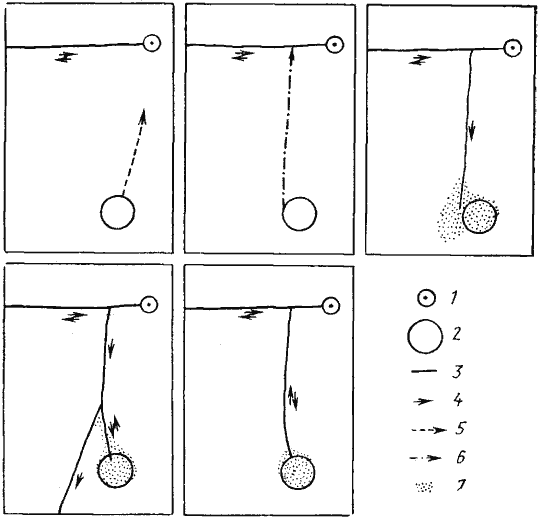
Рис. 88. Последовательные этапы формирования дороги к кормушке с пшеном фуражирами гнезда МА-18 Messor aralocaspius
(Акыбай, 30 апреля 1970 г.)
1 —гнездо; 2 — кормушка; 3 — дороги; 4 — направление движения пассивных фуражиров; 5 — путь первого разведчика; 6 — путь второго разведчика; 7 — зона рассева пассивных фуражиров. Размер закартированной площадки 1x1,2 м
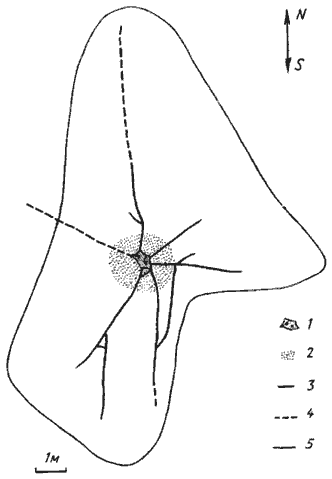
Рис. 89. Территория гнезда Messor denticulatus в семиаридных условиях
(Копетдаг, Ипай-Кала, № 71-127)
1 — пригнездовая очищенная площадка и входы в гнездо;
2 — участок с выбросами мякины;
3 — отчетливые заглубленные дороги;
4 — неявственные дороги;
5 — граница кормового участка 30 мая 1971 г.
Всего 106 видов.
(от греч. oikos дом, жилище, местопребывание и ...логия), наука об отношениях живых организмов и образуемых ими сообществ между собой и с окружающей средой. Термин «экология» предложен в 1866 Э. Геккелем. Объектами экологии могут быть популяции организмов, виды, сообщества, экосистемы и биосфера в целом.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
(гр. topos - место) - естественное жизненное пространство определенного биоценоза, совместно с которым он образует экосистему.
интенсивность посещения муравьями единицы площади в единицу времени (особь/дм²хмин.)
ксеробионт
гигрофил
участок, используемый муравьями для добычи пищи.
(от трофо… и греч. allaxis — обмен) передача пищи и гормональных веществ от особи к особи путем кормления содержимым зобика, желудка или слизывания выпота с поверхности тела. Играет огромную роль в передаче информации между муравьями.
часть гнезда рыжих лесных муравьев, занимаемая одной колонной.
пространство между охраняемыми территориями семей или колонн не посещаемое фуражирами из конкурирующих семей (колонн).
Старое название: Messor rufitarsis
(лат. mandibule — челюсть, от mando — жую, грызу), первая пара челюстей у ракообразных, многоножек и насекомых. Синоним: жвала.
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
организационно оформленная мультифункциональная часть семьи (подсистема), имеющая постоянный состав и способная перейти к самостоятельному существованию.
индивидуальное или групповое привлечение пассивных фуражиров активными фуражирами для конкретных полей в конкретном месте.
(греч. φέρω — «нести» + ορμόνη — «гормон») — летучие вещества, управляющие поведенческими реакциями, процессами развития, а также многими процессами, связанными с социальным поведением и размножением. Феромоны модифицируют поведение, физиологическое и эмоциональное состояние или метаболизм других особей того же вида.
Всего 60 видов.
Всего 172 вида.
Включает сокращенный Novomessor.
Старое название: Novomessor pergandei, Veromessor pergandei
ксеробионт






