






Прежде чем перейти к рассмотрению особенностей использования кормового участка у Cataglyphis, являющихся главными крупными зоонекрофагами пустынь Старого Света, необходимо кратко остановиться на экологии Alloformica aberrans. У этого вида, очень близкого к предкам Cataglyphis, наблюдается один из самых примитивных типов использования кормового участка. Радиус его составляет 5—10 м в местах со сравнительно обильной травянистой растительностью и 10—20 м на каменистых склонах со скудной растительностью. В пределах кормового участка все фуражиры работают как одиночные сборщики и контактов между ними (за исключением переселения в новые отводки) на территории ни разу не наблюдалось. Не заметно у этого муравья и разделения функций у внегнездовых рабочих. Одна и та же особь может собирать трупы насекомых или растительные остатки для закрывания входа в гнездо, кормиться на цветах и переносить других муравьев при переселении в новый отводок. Вторичное деление территории также отсутствует. Мы наблюдали, как одни и те же рабочие в два последующих дня кормились на цветах в разных секторах (к югу и к западу от гнезда) кормового участка. Видимо, как у ![]() Myrmica rubra (Длусский, 1975в) и Ponera coarctata, у них каждый фуражир имеет несколько предпочитаемых маршрутов. Каких-либо следов мобилизации и неспецифической активации в естественных условиях также не удалось обнаружить. Весной 1978 г. две семьи этого вида были привезены на Звенигородскую биостанцию и помещены в искусственные гнезда. В течение лета за ними велись наблюдения, но попытки вызвать неспецифическую активацию оказались безрезультатными. Рабочий, обнаруживший кормушку, наполнял зобик сиропом, уходил в гнездо и долгое время не появлялся на поверхности. В гнезде он либо кормил других муравьев, либо замирал в одной из камер. Возбужденного поведения в гнезде ни разу не наблюдалось. Опыты с белковыми приманками поставить было невозможно, так как к началу опытов цикл развития расплода уже закончился и в гнездах были только куколки.
Myrmica rubra (Длусский, 1975в) и Ponera coarctata, у них каждый фуражир имеет несколько предпочитаемых маршрутов. Каких-либо следов мобилизации и неспецифической активации в естественных условиях также не удалось обнаружить. Весной 1978 г. две семьи этого вида были привезены на Звенигородскую биостанцию и помещены в искусственные гнезда. В течение лета за ними велись наблюдения, но попытки вызвать неспецифическую активацию оказались безрезультатными. Рабочий, обнаруживший кормушку, наполнял зобик сиропом, уходил в гнездо и долгое время не появлялся на поверхности. В гнезде он либо кормил других муравьев, либо замирал в одной из камер. Возбужденного поведения в гнезде ни разу не наблюдалось. Опыты с белковыми приманками поставить было невозможно, так как к началу опытов цикл развития расплода уже закончился и в гнездах были только куколки.
В некоторых отношениях поведение Cataglyphis сходно с поведением A. aberrans, но в пределах этого рода мы можем наблюдать различные способы повышения эффективности использования кормового участка. Обычно участок имеет форму, близкую к кругу с радиусом около 10 м у Cataglyphis emeryi и C. pallida, 10—20 м — у C. cinnamomea и ![]() C. aenescens и 20—30 м — у C. foreli и C. setipes. У последнего вида иногда фуражиры уходят на расстояние до 60 м от гнезда. Ни у одного из видов рода весь кормовой участок не охраняется, но у C. foreli и C. setipes показано, что зона в непосредственной близости от гнезда охраняется от особей из других семей того же вида (Захаров, 1972). Взаимодействие фуражиров на территории в нормальных условиях отсутствует. Даже особи из одной семьи, встретившись вдали от гнезда, всегда обходят друг друга не приближаясь. Единственное исключение — совместная транспортировка крупной добычи двумя рабочими, изредка наблюдающаяся у C. pallida. Однако на кормушках с сахарным сиропом, особенно на расстоянии до 2 м от гнезда, муравьи могут кормиться большими группами.
C. aenescens и 20—30 м — у C. foreli и C. setipes. У последнего вида иногда фуражиры уходят на расстояние до 60 м от гнезда. Ни у одного из видов рода весь кормовой участок не охраняется, но у C. foreli и C. setipes показано, что зона в непосредственной близости от гнезда охраняется от особей из других семей того же вида (Захаров, 1972). Взаимодействие фуражиров на территории в нормальных условиях отсутствует. Даже особи из одной семьи, встретившись вдали от гнезда, всегда обходят друг друга не приближаясь. Единственное исключение — совместная транспортировка крупной добычи двумя рабочими, изредка наблюдающаяся у C. pallida. Однако на кормушках с сахарным сиропом, особенно на расстоянии до 2 м от гнезда, муравьи могут кормиться большими группами.
У C. foreli имеется хорошо выраженное вторичное деление территории (Захаров, 1972), которое носит секторный характер. Каждый фуражир изо дня в день работает на индивидуальном поисковом участке в пределах сектора. Площадь индивидуального участка достигает 20 м². Охотники, действующие на периферии кормового участка, не трогают добычи, находящейся неподалеку от гнезд. Сходная картина наблюдается и у C. setipes. Как было показано А. А. Захаровым (1975а) на примере C. setipes, поведение этих муравьев во многом зависит от плотности особей на кормовом участке. При искусственном увеличении динамической плотности с 0,14—0,31 особей/дм²/мин (до начала опыта) до 4—5 особей/дм²/мин (часть территории с гнездом была огорожена стенкой) муравьи начали проявлять вне гнезда некоторые элементы внутригнездового поведения. Они стали агрессивными по отношению к другим муравьям, начали коллективно нападать на более крупных животных (чернотелок, жужелиц) и коллективно транспортировать крупную добычу в гнездо. В нормальных условиях такое поведение на кормовом участке характерно для видов с более высоким уровнем социальной организации, например для Formica s. str.
Cataglyphis отличаются хорошим зрением. При движении на территории они руководствуются главным образом визуальными ориентирами — солнцем, поляризацией неба и наземными вехами (Burkhalter, 1972; Wehner, Flatt, 1972; Duelli, Wehner, 1973; Duelli, 1976; Францевич и др., 1976). Кроме того, для ориентации при слабой освещенности они могут использовать направление ветра (Duelli, 1972). Обычно поисковый рейд фуражиров состоит из пробежек длиной 1—2 м по прямой или по широкой дуге, чередующихся с остановками для ориентировки. Особенно характерно ориентировочное поведение C. foreli и C. emeryi, которые после пробежек останавливаются и делают оборот на 360°. Иногда муравьи для ориентировки поднимаются на травинки и другие возвышающиеся предметы.
Наиболее заметные отличия, наблюдаемые у разных видов Cataglyphis, связаны с различиями реакций этих муравьев на появление источника обильной пищи. У C. emeryi, самого примитивного морфологически представителя рода, имеющего самые малочисленные семьи (до 500 особей), в естественных условиях нам не удалось наблюдать ни мобилизации, ни неспецифической активации. Плотность особей вне гнезд у этого вида очень низка. Для выяснения потенциальных возможностей C. emeryi одна семья летом 1974 г. была привезена из Карши и помещена в лабораторное гнездо на Звенигородской биостанции МГУ. Гнездо соединялось с ареной размером 120X40 см, в пределах которой стеклянными перегородками было выделено три сектора (А, Б и В) приблизительно одинаковой площади и предгнездовая площадка (ГШ) размером 20X25 см (рис:69). Семья состояла из 300 рабочих, самки и некоторого количества расплода, главным образом куколок. Заселение проведено 20 июня, а первые опыты начались месяц спустя, когда муравьи освоились с искусственным гнездом и ареной. Все это время корм давался в секторах А и В. 18 и 19 июля в секторе А было промечено индивидуальными метками 15 рабочих. В последующие дни девять из них работали на арене, в том числе 5 — в том же секторе, один — в секторах Б и В, один — в секторе В и один — в А и Б. Рабочий, промеченный в секторе В, в последующие дни работал здесь же.
Первый опыт был поставлен 20 июля (табл.27). Вначале подсчитывалось количество рабочих, входящих в каждый сектор в течение пятиминутных интервалов (контроль). Затем в сектор В была поставлена кормушка. Отсчеты начинались после обнаружения корма первым фуражиром (опыт). Хотя некоторые возвращавшиеся от кормушки муравьи выглядели явно возбужденными и активно бегали по гнезду, реакции семьи на появление корма не последовало. Ни в одном из секторов число проходов достоверно не увеличилось. После этого семья в течение четырех дней не получала пищи и 24 июля опыт был повторен. Кормушка также была поставлена в сектор В. В этом опыте число проходов в сектор В достоверно возросло (Р<0,01), а в секторах А и Б не изменилось по сравнению с контролем (табл.26, рис:69). Однако среднее число муравьев, одновременно находящихся в секторе, достоверно не изменилось ни в одном из секторов. В первые полчаса после того, как муравьи начали возвращаться в гнездо, на пригнездовой площадке наблюдалось увеличение числа муравьев (Р=0,05), что указывает на слабую активацию. Прямые наблюдения показали, что активация действительно произошла, но большинство вышедших из гнезда муравьев возвращались обратно, а доставку корма производили одни и те же несколько фуражиров, случайно обнаруживших кормушку. При последующих кормлениях картина в общем повторялась, т.е. в тех случаях, когда семья получала пищу регулярно, приход фуражиров с кормушки не вызывал заметного эффекта. Если же муравьи долго не получали пищи, некоторое количество активированных рабочих выходило на пригнездовую площадку, но число муравьев в секторах заметно не увеличивалось.
Таким образом, у C. emeryi, как и у A. aberrans, в нормальных условиях семья не реагирует на увеличение количества пищи на территории. Даже в том случае, когда муравьи долго не получают пищи, кормушка выставляется в освоенном секторе и близко от гнезда (в опытах — 80 см) наблюдается лишь очень слабая активация семьи, не приводящая к увеличению числа фуражиров на территории. На основании наших опытов можно также предположить, что у C. emeryi имеется вторичное деление территории, поскольку даже в условиях, когда искусственная арена была в несколько десятков раз меньше естественной территории, наблюдалась явная привязанность рабочих к секторам.
Для выяснения способностей C. setipes к неспецифической активации в Акыбае в марте 1970 г. была проведена серия опытов на естественных гнездах. В одной из серий (26—30 марта, гнездо КС-701) в первый день на расстоянии 80 см от гнезда была поставлена кормушка с сахарным сиропом. Все приходившие на нее муравьи метились нитролаками индивидуальными метками. Через 20 мин после начала опыта (рис:70) на кормушке работали четыре муравья (№1—4). После того, как эти фуражиры сделали по два рейса в гнездо, из него вышла новая группа рабочих, три из которых (№6—8) пришли на кормушку. Через 2 ч кормушку убрали. В это время на ней находились семь особей. На следующий день кормушка была выставлена в то же место. Вначале на ней работали главным образом рабочие, посещавшие ее во время прошлого опыта (№1, 3, 5, 6 и новый №9). Через час на ней появилась новая группа особей, среди которых только два (№2, 4) работали раньше. Общая численность муравьев, находящихся одновременно на кормушке, возросла до десяти. 28 марта корм не выставлялся, а 29 он был выставлен на 4 ч 30 мин на том же месте. На этот раз вначале работала группа из четырех новых (№15—18) и одного старого (№3) рабочих. Через 30—50 мин после начала опыта вышла первая группа активированных рабочих (все новые), а через 2,5 ч — вторая (также все новые). Кроме того, в течение первых 2,5 ч опыта поодиночке на корме появились пять рабочих, участвовавших в опытах 26 и 27 марта. Общее число муравьев, одновременно находящихся на кормушке, возросло к концу опыта до 14.
30 марта была выставлена новая кормушка, расположенная в 80 см от гнезда и от места прежнего кормления. Предварительно в течение часа каждые 5 мин подсчитывалось число муравьев, находящихся в квадрате 20X20 см. В квадрате на месте будущей кормушки оно было равно в среднем 2,0±0,8 особей, а в квадрате, выставленном на том месте, где муравьи получали корм в предшествующих опытах, — 9,0±0,8 особей. Через 40 мин после того как кормушка была поставлена, на ней одновременно находились до 11 особей, а к концу опыта — 14. На месте прежнего кормления в квадрате 20X20 см в течение первых двух часов опыта в среднем одновременно находились 3,0±0,7 особей, а к концу опыта — 1,0±0,3 особей (рис:70). Таким образом, результаты этих опытов показывают, что рабочие C. setipes, во всяком случае при выставлении кормушки в пригнездовой зоне, могут вызывать в гнезде неспецифическую активацию, в результате которой из гнезда выходит от трех до пяти активированных муравьев.
Неспецифическую активацию у C. setipes можно наблюдать и в естественных условиях. Так, в приведенном выше (рис:38) опыте по изучению суточной активности этого вида в первой половине дня наблюдались два пика интенсивности выхода из гнезда: первый — после возвращения в гнездо с добычей первых фуражиров и второй — после прихода муравьев с трупами муравьев-жнецов. В жизни семьи C. setipes неспецифическая активация играет важную роль прежде всего при сборе трупов Messor aralocaspius. Гнезда этих двух видов часто располагаются неподалеку, но во время фуражировки муравьев-жнецов фаэтончики не заходят на их территорию. В то время, когда большинство пассивных фуражиров муравьев-жнецов занимается сбором семян, некоторые особи чистят гнездо и выносят по тем же дорогам различный мусор и трупы погибших в гнезде муравьев. Как только температура на поверхности почвы поднимается до 38—40°, все рабочие муравьев-жнецов скрываются в гнездах. И тогда на их участки устремляются фаэтончики, которым необходимо быстро собрать все трупы, пока их не унесли другие муравьи. Настоящая мобилизация в этой ситуации не имеет смысла, так как трупы рассеяны на значительной площади. Такие же взаимоотношения мы наблюдали у ![]() C. aenescens и M. denticulatus (Длусский, 1975а). Неспецифическая активация также наблюдалась у C. pallida, значительную долю в питании которого составляют трупы M. aralocaspius и Tetramorium schneideri. Важное значение имеет неспецифическая активация во время брачного лёта термитов и муравьев. Крылатые половые особи термитов и других видов муравьев — излюбленная добыча всех средних и крупных зоонекрофагов.
C. aenescens и M. denticulatus (Длусский, 1975а). Неспецифическая активация также наблюдалась у C. pallida, значительную долю в питании которого составляют трупы M. aralocaspius и Tetramorium schneideri. Важное значение имеет неспецифическая активация во время брачного лёта термитов и муравьев. Крылатые половые особи термитов и других видов муравьев — излюбленная добыча всех средних и крупных зоонекрофагов.
У C. foreli при постановке опытов, аналогичных описанным выше, мы не наблюдали неспецифической активации. Так, например, 5 апреля 1970 г. в Каррыкуле была выставлена кормушка с сахарным сиропом в 80 см от гнезда КФ-701 (рис:71). Как и в опытах с C. setipes, все фуражиры, обнаружившие кормушку, метились индивидуальными метками. Опыт начался в 11 ч, когда первый фуражир был замечен на кормушке. До 13 ч 50 мин на кормушку вышли еще четыре муравья, каждый из которых находил ее самостоятельно. В последующие 2 ч появился еще один рабочий. С 17 до 19 ч на кормушке работали четыре фуражира и новых муравьев больше не появлялось.
Иная картина наблюдается в том случае, если C. foreli предложить трупы термитов — излюбленной пищи этих муравьев. 6 апреля 1970 г. в Каррыкуле в 2,2 м от гнезда КФ-702 была выложена кучка свежеубитых термитов. Первый рабочий обнаружил приманку в 11 ч 15 мин и от этого момента начали отсчет по секундомеру (рис:71/2/). Через 30 с этот фуражир вошел в гнездо с добычей и еще через 40 с вышел из гнезда. Вместе с ним вышли еще две особи. До этого муравьев возле входа не было, а с этого момента на пригнездовой площадке постоянно находились одна — пять особей. Муравьи то выбегали из гнезда, то скрывались в нем. В 11 ч 19 мин 55 с разведчик вернулся со вторым термитом и в последующие 30 с из гнезда вышла новая группа рабочих. Общее число особей, одновременно находящихся на площадке вокруг входа, возросла до 6—10. К моменту третьего возвращения (11 ч 21 мин 55 с) эта группа образовала подобие клина, направленного острием к кормушке. Одновременно часть вышедших муравьев начала уходить в гнездо. К 11 ч 22 мин 50 с на поверхности осталось только два муравья, которые стояли в 30 см от входа в гнездо на прямой, соединяющей его с кормушкой. Время от времени они делали небольшие (менее 90°) повороты всем телом, но общая ориентация на кормушку в среднем сохранялась. После того как разведчик ушел в четвертый рейс, они поодиночке направились прямо к кормушке и начали также носить термитов в гнездо. Сходная картина наблюдалась и в других опытах. На рис:71/3/ показаны некоторые результаты аналогичного опыта, поставленного в тот же день на гнезде КФ-703. Основное отличие заключалось в том, что активировано было более 30 рабочих (семья была большая). Мобилизовались три рабочих после 3-, 6- и 8-го рейсов разведчика и соответственно через 10 мин, 20 мин 20 с и 29 мин после первого возвращения разведчика в гнездо. В нескольких опытах разведчик изымался после 1—2-го рейсов. Мобилизации при этом никогда не происходило, а активированные рабочие (если такие были) уходили в гнездо.
Наблюдавшийся процесс не был обычной мобилизацией за счет движения тандемом или привода. В опыте на гнезде КФ-702, например, муравей №2 пошел тогда, когда разведчик (№1) находился уже на кормушке, а №3 — когда №1 уже вернулся с добычей и скрылся в гнезде, а №2 находился около кормушки. Вообще, очень часто мобилизованные особи начинали двигаться через несколько секунд после того, как мимо них проходил в гнездо рабочий, несущий добычу (разведчик или один из мобилизованных). В то же время это нельзя назвать и неспецифической активацией, так как муравьи сразу идут в направлении кормушки и почти не тратят времени на поиск. Использование пахучего следа, нанесенного на субстрат, невозможно, так как все Cataglyphis бегут с большой скоростью, высоко подняв брюшко над землей. Сигнализация пахучим следом в воздухе, наличие которого у Catagluphis предполагал Гетч (Goetsch, 1953), также невозможна, так как большинство опытов с C. foreli проводилось на лишенных растительности площадках на склонах барханов во время ветреной погоды. Все поведение муравьев во время мобилизации указывает, что активированные фуражиры выходят из гнезда и смотрят, куда и откуда ходит разведчик, приносящий добычу. После того как он совершит два-три рейса у них на виду, активированные особи идут в том же направлении и находят кормушку. Такую форму мобилизации, не описанную ранее ни у одного из муравьев, можно назвать киноптической мобилизацией. В отличие от мобилизации путем обычного тандема или пахучего следа, для ее осуществления требуется высокая степень развития пространственной ориентации и развитая кратковременная память. Появление киноптической мобилизации у C. foreli явно связано с пищевой специализацией этого муравья. Излюбленной пищей этих муравьев являются термиты Anacanthotermes ahngerianus. Однако фуражиры C. foreli активны исключительно днем, когда термиты не выходят из лепных тоннелей. Стоит разрушить такой тоннель или повредить гнездовой купол, как очень быстро появляется несколько рабочих C. foreli, которые начинают охотиться на бегающих вокруг повреждения термитов. Обычно им при этом удается унести не более десятка термитов, так как последние быстро скрываются в тоннеле. В таких условиях оптимальной стратегией будет быстрая мобилизация немногих особей, которую и обеспечивает киноптическая мобилизация.
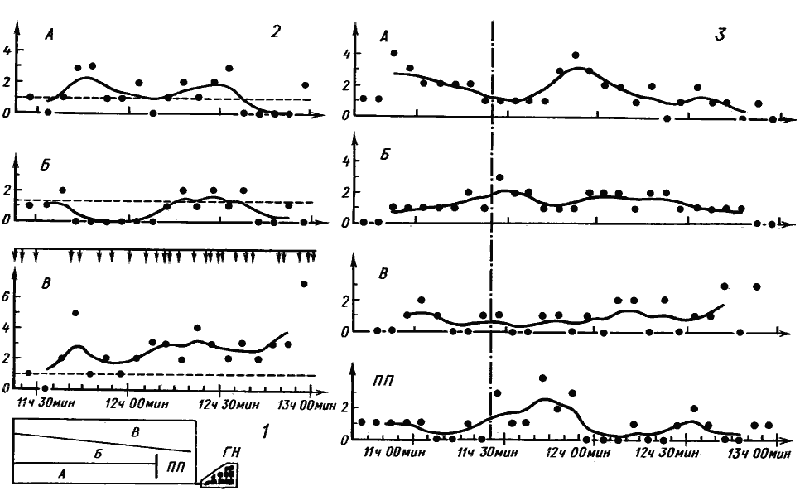
Рис. 69. Реакция семьи Cataglyphis emeryi на кормушку с сахарным сиропом
(лабораторный эксперимент 24 июля 1974 г.)
1 — схема установки: ГН — гнездо, ПП — пригнездовая площадка; А, Б, В — субарены; размеры арены 120X40 см; кормушка поставлена в субарену В в 10 ч 15 мин; первый фуражир с кормом вышел из субарены В в 10 ч 25 мин; 2 — число проходов в субарены за 5 мин (точки) в течение опыта, кривая получена в результате обработки данных методом скользящей кривой; пунктир — среднее число проходов в сектор за 5 мин в контроле. Вертикальными стрелками показано время выходов фуражиров с кормом из субарены В; 3 — число рабочих, находившихся в субаренах и на пригнездовой площадке, в контроле (до 10 ч 25 мин) и в опыте
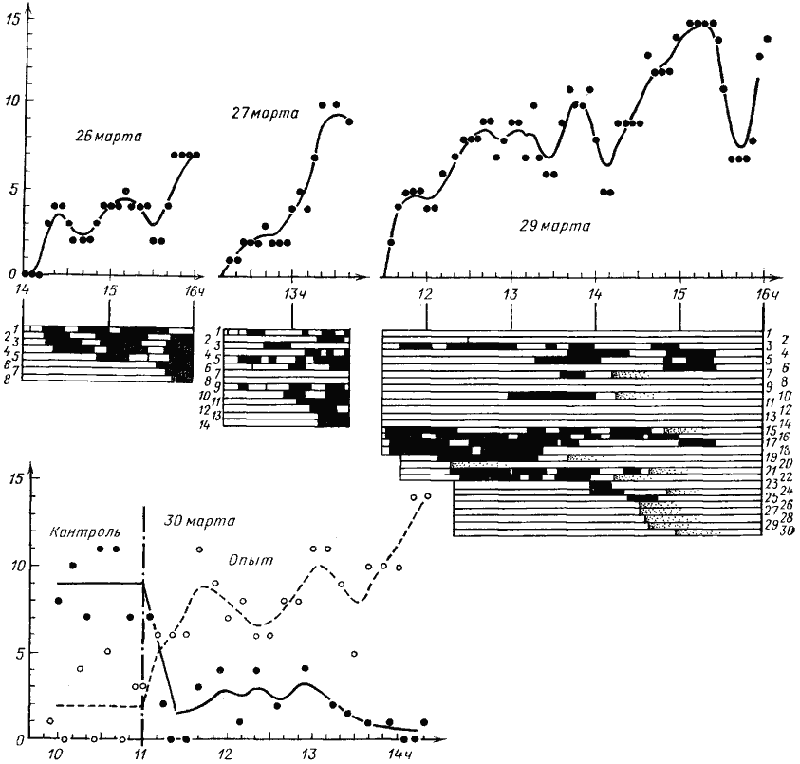
Рис. 70. Реакция семьи Cataglyphis setipes на кормушку с сахарным сиропом, расположенную вблизи гнезда
(Акыбай, № КС-701)
26 марта с 14 до 16 ч была выставлена кормушка с сахарным сиропом на расстоянии 1 м от гнезда. 27 и 29 марта кормушка выставлялась на том же месте, соответственно с 12 ч 10 мин до 13 ч 40 мин и с 11 ч 30 мин до 16 ч. Все особи, появившиеся на кормушке, метились индивидуальными метками и им присваивался порядковый номер. На верхних графиках показано изменение числа особей на кормушке (отметки через 5 мин и усредненная кривая), под ними — время пребывания на кормушке отдельных особей (зачернено). 29 марта в ряде случаев (например, особи № 20, 26—30) отмечался только момент появления на кормушке и дальше наблюдения за этой особью не велись. 30 марта с 9 ч 30 мин до 11 ч отмечалось число особей на площадке, где в предыдущие дни выставлялась кормушка (зачерненные кружки, сплошная линия), и на такой же площадке в 1 м от гнезда и в 1 м от бывшей кормушки (контурные кружки и пунктир). В 11 ч на второй площадке была выставлена кормушка и каждые 5—10 мин подсчитывалось число особей на обеих площадках
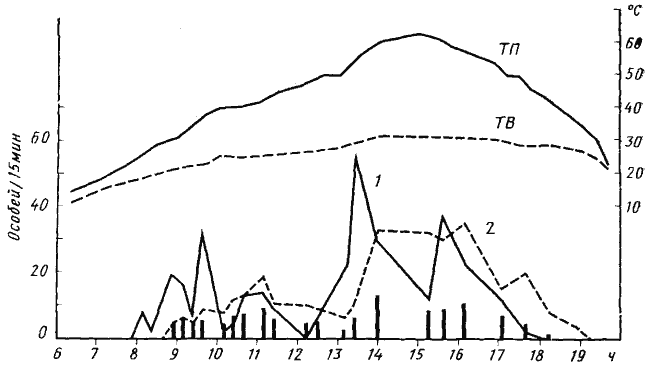
Рис. 38. Суточная динамика фуражировочной активности Cataglyphis setipes
(Акыбай. 10 мая 1971 г., гнездо №71-30)
ТВ — температура воздуха в тени на высоте 1 м; ТП — температура поверхности почвы на солнце (до 13 ч 15 мин вход в гнездо находился в тени саксаула и температура на поверхности почвы вблизи него была приблизительно равна температуре воздуха в тени);
1 — интенсивность выходов из гнезда (выходов/15 мин); 2 — интенсивность возвращения в гнездо; вертикальные столбики — число единиц добычи, принесенных в гнездо за 15 мин
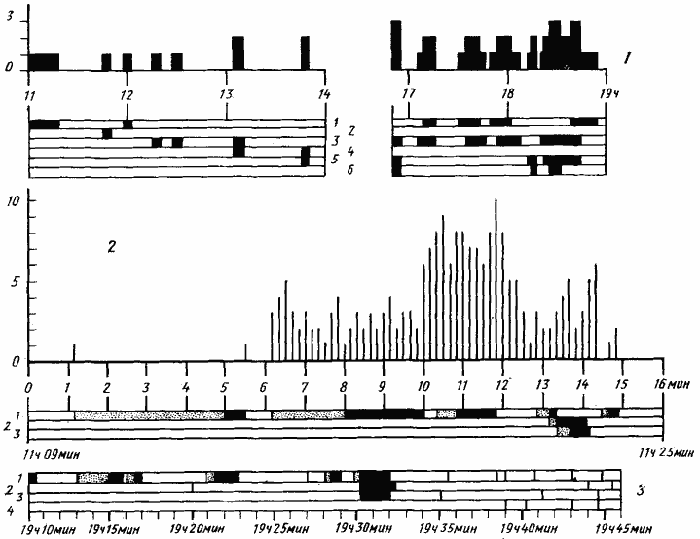
Рис. 71. Реакция Cataglyphis foreli на кормушки
(Каррыкуль, 1970 г.)
1 — кормушка с сахарным сиропом была выставлена 5 апреля в I м от гнезда КФ-701. Вверху — число особей на кормушке, внизу — время пребывания на кормушке шести кормившихся на ней фуражиров; 2 — кормушка с трупами термитов выставлена в 3 м от гнезда КФ-702 6 апреля. Вверху — число муравьев в пригнездовой зоне (вертикальные линии), внизу — этограммы рабочих, посещавших кормушку (точка — поиск добычи, зачернено — транспортировка добычи в гнездо); 3 — кормушка с трупами термитов выставлена в 1,5 м от гнезда КФ-703 6 апреля. Этограммы фуражиров, посещавших кормушку. В ряде случаев отмечался только момент прихода муравья на кормушку (вертикальная линия)
участок, используемый муравьями для добычи пищи.
Всего 65 видов.
(зоонекрофаг), организм, питающийся мертвыми животными, гетеротроф
(от греч. oikos дом, жилище, местопребывание и ...логия), наука об отношениях живых организмов и образуемых ими сообществ между собой и с окружающей средой. Термин «экология» предложен в 1866 Э. Геккелем. Объектами экологии могут быть популяции организмов, виды, сообщества, экосистемы и биосфера в целом.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
дочерний муравейник: гнездо, в котором обитает дочерняя семья (в колонии)
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
подразделение кормового участка семьи на элементарные поисковые участки, обслуживаемые одним фуражиром или небольшими группами
часть гнезда рыжих лесных муравьев, занимаемая одной колонной.
 Myrmica rubra (Linnaeus, 1758) — рыжая мирмика
Myrmica rubra (Linnaeus, 1758) — рыжая мирмикаиндивидуальное или групповое привлечение пассивных фуражиров активными фуражирами для конкретных полей в конкретном месте.
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
ксеробионт
Старое название: Proformica deserta
Старое название: Cataglyphis cinnamomea
 Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунок
Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунокгигрофил, галобионт
Старое название: Cataglyphis flavigastra
ксеробионт
ксеробионт
интенсивность посещения муравьями единицы площади в единицу времени (особь/дм²хмин.)
ксеробионт
гигрофил
отряд общественных насекомых. Общины, разделенные на касты, состоят из крылатых и бескрылых особей. Строят подземные и наземные (до 15 м высотой) гнезда (термитники). Ок. 2600 видов, главным образом в тропиках; в России 2 вида: один в районе Сочи, второй, вероятно, завезенный из Японии или Китая, во Владивостоке. Многоядны. Разрушают древесину, портят бумагу, кожу, сельскохозяйственные продукты и др. Термиты, обитающие в почве, полезны как почвообразователи.






