






Как уже говорилось в начале главы, жизнь в подземных гнездах обеспечила муравьям возможность существования в аридных условиях. Их личинки, не способные переносить высыхания, постоянно находились в почве, где даже в пустыне можно подобрать условия температуры и влажности, близкие к оптимальным. Однако освоение пустынь требовало дальнейшего совершенствования гнездостроения. И, естественно, гнезда пустынных муравьев обладают рядом черт, отличающих их от гнезд видов гумидных областей. Следует подчеркнуть, что адаптация к жизни в почвах пустынь шла у муравьев главным образом по пути усовершенствования строения гнезд, а не за счет изменения поведения при рытье почвы или появления специализированных морфологических структур. Действительно, псаммофоры муравьев, ставшие благодаря работам классиков мирмекологии хрестоматийным примером морфологических адаптации к жизни в пустынях, — всего лишь удлиненные и утолщенные фиксированные щетинки и (у некоторых видов) удлиненные нижнечелюстные щупики. Эти изменения не идут ни в какое сравнение с морфологическими изменениями одиночных животных, обитающих в почвах пустынь, таких, как тараканы Polyphagidae, клопы Stebaropus или многие чернотелки и хрущи. Однако эти сравнительно небольшие изменения позволили муравьям повысить эффективность рытья и за счет этого строить более глубокие или более диффузные гнезда.
Основным направлением прогрессивной (ароморфной) эволюции общественных насекомых, и в частности муравьев, является увеличение размеров семьи. Только в большой семье возможен развитой полиэтизм (разделение функций между рабочими), создание значительных запасов пищи, активная регуляция микроклимата в гнезде и т.д. В конечном счете увеличение числа особей в семье обеспечивает биологический прогресс социальных насекомых, повышая гомеостатичность семьи и стабильность размножения семей. С увеличением размеров семьи неразрывно связано увеличение размеров гнезда (за исключением муравьев-кочевников) и усложнение его структуры. Эти процессы происходили и происходят независимо во многих группах социальных насекомых. Однако, поскольку исходная база (примитивное гнездо и примитивная структура семьи) примерно одинакова в разных исходных группах муравьев, а среда задает сходные ограничения, мы можем наблюдать параллельное возникновение сходных структур семьи и сходных типов сложных гнезд. Наряду с изменениями структуры гнезд, связанными с увеличением размеров семьи, в каждой природной зоне мы наблюдаем возникновение теломорфных или алломорфных приспособлений, дающих преимущества в условиях данной природной зоны. В лесах умеренной зоны развиваются, например, различные способы создания температурного режима, благоприятного для развития расплода (Длусский, 1967а, 19726). В аридных условиях возникают специфические особенности структуры гнезд, позволяющие создать благоприятные для развития расплода условия влажности.
Первое, что бросается в глаза при анализе гнездостроения муравьев в пустынях, — это разделение гнезд на три довольно четкие группы, различающиеся по глубине проникновения стволов (табл.24). К первой группе относятся все мелкие муравьи (средний сухой вес рабочего менее 0,5 мг), за исключением Monomorium barbatulum и Acantholepis semenovi. Глубина их гнезд редко достигает 60—70 см и обычно составляет 30—50 см. Все эти виды используют для жизни поверхностный горизонт почвы, в котором весной и осенью имеется достаточно влаги. Некоторые из этих видов обитают только во влажных биотопах. У Plagiolepis, Tetramorium inerme и M. barbatulum имеется сеть тонких галерей, идущих под самой поверхностью, иногда на глубине нескольких миллиметров (рис:65/3/; рис:68/3/). Возможно, эти муравьи способны использовать воду, конденсирующуюся ночью на поверхности почвы, как это делают, например, многие кактусы. В первой половине весны расплод в таких гнездах находится на глубине до 20 см, но к началу лета самые верхние слои пересыхают и муравьи опускают личинок в камеры на глубине 40—50 см. Интересно, что Cataglyphis (рис:60/1/) и Acantholepis весной также иногда используют только поверхностный горизонт, а глубинные ходы таких гнезд оказываются брошенными и частично засыпанными почвой. То же описано для C. bicolor в Сахаре (Delye, 1968).
Вторая группа видов, куда относятся все пустынные Cataglyphis, Monomorium barbatulum, Acantholepis semenovi, Messor variabilis и, возможно, M. excursionis и M. lamellicornis, использует глубинный горизонт (глубже 70—100 см), где в течение всего года влажность почвы остается выше МГ. За исключением M. barbatulum и A. semenovi, все это сравнительно крупные муравьи (средний сухой вес рабочих 0,5—7,5 мг), имеющие более или менее развитый псаммофор. M. barbatulum имеет мелкие размеры, но его псаммофор развит необычайно сильно. A. semenovi обычно не строит гнезд самостоятельно, а использует брошенные гнезда термитов или крупных муравьев и лишь слегка перестраивает их или поселяется в трещинах скал. В гнездах представителей этой группы муравьев расплод находится либо в ПГС, либо в самых глубоких камерах гнезда.
Представители третьей группы, куда относятся Camponotus xerxes, C. turcestanicus, Messor denticulatus, M. aralocaspius, M. intermedius и, возможно, другие виды Messor и Camponotus, строят гнезда, стволы которых достигают грунтовой воды. Всё это крупные муравьи (средний сухой вес рабочих 2,2—19,4 мг) с хорошо развитым псаммофором. Они (особенно Camponotus) охотно пьют капельную воду. Характерно и стациальное распределение видов этой группы. Одни из них в условиях пустынь Средней Азии встречаются исключительно на солончаках, в тугаях или на орошаемых землях (M. denticulatus, C. turcestanicus). В пустыню они заходят лишь в зоне, граничащей с оазисами, где глубина залегания грунтовых вод не превышает 4—5 м. В последнем случае большинство камер гнезда сгруппировано в глубине, над самым водоносным горизонтом (табл.25). Другие виды (C. xerxes, M. aralocaspius) обитают преимущественно в пустынях, так как в оазисах и тугаях они вытесняются гигрофильными видами (Длусский, 1963; Сапарлиев, 1971). Наибольшей численности эти муравьи достигают в участках пустыни, граничащих с оазисами, но могут быть обнаружены в центре Каракумов или Кызылкумов. В Центральных Каракумах гнезда этих видов располагались исключительно по краям такыров, на которых грунтовые воды подходили сравнительно близко к поверхности. Таким образом, представители этой группы муравьев могут, как справедливо указывал П. И. Мариковский (1957) для M. aralocaspius, служить индикатором грунтовых вод в пустынях. В глубоких гнездах этих муравьев весной основная масса расплода находится в поверхностном горизонте и в узкой зоне на глубине между 1,5 и 2,5 м. В качестве примера можно привести распределение рабочих и расплода в гнезде молодой семьи C. xerxes глубиной 390 см (табл.26).
Возможно, исходным для некоторых групп муравьев, все формирование которых шло в условиях пустыни, было сфекоидное гнездо, сходное с таковым у Alloformica. Во всяком случае, это утверждение, видимо, справедливо для Cataglyphis, происхождение которых от Alloformica не вызывает сомнения. Даже такое гнездо позволяет муравьям использовать градиент влажности в почве. Но оно имеет существенный недостаток: такое гнездо, фактически представляющее собой вертикальную шахту, плохо удерживает влажный воздух. При этом пропитка стенок хода секретами желез только ухудшила бы положение, так как замедляла бы испарение влаги из почвы в гнездо. У пустынных мокриц, норки которых очень похожи на гнезда A. aberrans, одна из взрослых особей постоянно сидит у входа, закрывая его своим телом.
Первыми шагами на пути повышения экономии влаги было появление камер и удаление входа в гнездо от вертикального ствола. Конечно, появление камер у гнезда муравьев нельзя связывать с адаптацией к аридности, так как они хорошо развиты в гнездах большинства видов гумидной зоны. Камеры, очевидно, возникали как приспособление, связанное с увеличением размера семьи и увеличением количества расплода. Однако побочным эффектом было повышение экономии влаги: камера связана с вертикальным стволом только узким проходом. Кроме того, появление камер позволило сузить сам проход. Немаловажное значение также имело то обстоятельство, что муравьи уплотняли стенки ходов и камер и покрывали их секретами желез. Поверхностная горизонтальная галерея, соединяющая вход с гнездом и не имеющая камер, встречается только у пустынных муравьев. Вход может быть удален от основного гнезда на расстояние около 1 м, как это наблюдается у Monomorium chobauti (Delye, 1968) или Cataglyphis cinnamomea (рис:56). Крайней степени стремление избежать потерь воды и в то же время использовать преимущества верхнего горизонта (повышенный градиент температуры и высокая влажность после дождей) достигает в гнездах с функциональной дифференциацией стволов (Messor variabilis, рис:63) или секций (Camponotus xerxes; рис:67; Monomorium barbatulum, рис:68) гнезда. Расплод размещается в специальном отделе, не имеющем самостоятельного выхода (и соответственно отверстия, через которое происходит повышенное испарение влаги) и соединяющемся с остальным гнездом лишь тонкой горизонтальной галереей, часто проходящей на большой глубине. Такие гнезда имеются также только у муравьев пустынь и являются теломорфной адаптацией к специфическим условиям аридной зоны.
Главной линией ароморфной эволюции структуры гнезд, сопряженной с увеличением размеров семьи, была линия гнездо с одним стволом без ПГС → гнездо с одним стволом и развитой ПГС → гнездо с несколькими стволами и ПГС → многосекционное гнездо. По этой базовой схеме эволюция шла независимо во многих группах муравьев как в аридной зоне, так и в гумидных областях с умеренным климатом. На это указывает тот факт, что в пределах родственной группы видов определенному уровню развития социальности соответствует определенный тип гнезда.
Так, например, в окрестностях Звенигорода (Московская область) у гнезд Myrmica schencki (численность населения не более 1000 особей, обычно 300—500) ПГС редко бывает развита, а для ![]() M. rubra (до 10 тыс. особей) характерно гнездо с развитой ПГС и одним стволом. В Воронежском заповеднике у
M. rubra (до 10 тыс. особей) характерно гнездо с развитой ПГС и одним стволом. В Воронежском заповеднике у ![]() Formica fusca (население до 1000 особей) мы находили чаще всего гнезда без развитой ПГС, у
Formica fusca (население до 1000 особей) мы находили чаще всего гнезда без развитой ПГС, у ![]() F. cunicularia (1—10 тыс. особей) — с развитой ПГС и одним стволом и у F. imitans (до 100 тыс., а возможно и более) — односекционные гнезда с несколькими стволами или многосекционные. Интересно отметить, что в онтогенезе гнездо также проходит эти стадии. Так, например, недавно основанные гнезда Camponotus xerxes состоят из одного ствола и не имеют ПГС, семьи, численность которых достигает 500—600 особей, имеют гнездо с ПГС и одним стволом (рис:67/1/), семьи с численностью 1,5—2 тыс. особей — гнездо с двумя-тремя стволами, а численностью более 3—4 тыс. — многосекционное гнездо (рис:67/2/). Примерно такая же картина наблюдается у Tetramorium schneideri или Formica imitans.
F. cunicularia (1—10 тыс. особей) — с развитой ПГС и одним стволом и у F. imitans (до 100 тыс., а возможно и более) — односекционные гнезда с несколькими стволами или многосекционные. Интересно отметить, что в онтогенезе гнездо также проходит эти стадии. Так, например, недавно основанные гнезда Camponotus xerxes состоят из одного ствола и не имеют ПГС, семьи, численность которых достигает 500—600 особей, имеют гнездо с ПГС и одним стволом (рис:67/1/), семьи с численностью 1,5—2 тыс. особей — гнездо с двумя-тремя стволами, а численностью более 3—4 тыс. — многосекционное гнездо (рис:67/2/). Примерно такая же картина наблюдается у Tetramorium schneideri или Formica imitans.
Выше уже отмечалось, что во всех типах гнезд пустынных муравьев значительная часть расплода находится в верхних камерах гнезда, как и у муравьев гумидных областей умеренной зоны. Это объясняется тем, что в ряде отношений поверхностный слой почвы является наиболее благоприятным для развития муравьев. Пока большую часть расплода семьи можно разместить в одной-двух камерах, гнездо с одним стволом без ПГС вполне удовлетворяет потребности семьи. Фактически такое гнездо полностью аналогично термограду Гертера, поставленному вертикально. Однако при увеличении размеров семьи число камер увеличивается главным образом в верхних горизонтах гнезда. Глубинные камеры могут располагаться друг над другом, так как градиент микроклимата на глубине меняется не так резко. Лишь в пустыне могут возникать глубинные горизонтальные системы в глубоком влажном слое почвы.
При дальнейшем увеличении размеров семьи одного ствола может оказаться мало. Поскольку в аридных условиях у видов, использующих глубинные влажные горизонты почвы, нормальные функции может выполнять только полностью готовый ствол (т.е. идущий по крайней мере до глубины порядка 1 м), усложнение структуры гнезда идет скачкообразно. В гумидных районах эта картина может быть смазанной. Так, у ![]() Lasius flavus увеличение размера гнезда идет главным образом за счет ПГС, располагающейся в земляной кочке. Даже у самых сильных семей этого вида гнездо имеет только один ствол, правда, с очень крупными камерами, в которые вся семья может собираться для зимовки.
Lasius flavus увеличение размера гнезда идет главным образом за счет ПГС, располагающейся в земляной кочке. Даже у самых сильных семей этого вида гнездо имеет только один ствол, правда, с очень крупными камерами, в которые вся семья может собираться для зимовки.
В процессе эволюции увеличение размеров семьи в конце концов приводит к тому, что семья перестает быть монолитным целым и распадается на структурные субъединицы — колонны. Связь между особями в пределах колонны гораздо выше, чем между колоннами, хотя обмен рабочими, расплодом и пищей происходит. Каждая колонна имеет свой кормовой участок (Захаров, 1972, 1976а). С формированием колонн связано появление многосекционного гнезда. В большинстве случаев секции большого гнезда располагаются довольно компактно и ПГС разных секций соприкасаются. Исключение представляет уникальное диффузное гнездо Monomorium barbatulum, секции которого удалены друг от друга. Такое гнездо занимает большую площадь и, помимо выполнения основных, присущих всем гнездам муравьев, функций, обеспечивает M. barbatulum возможность существования в условиях крайне скудной кормовой базы. Этот муравей обитает на участках, большую часть года лишенных растительности, где другие мелкие карпофаги существовать не могут. Радиус действия фуражиров этого крошечного муравья редко достигает 2 м, а обычно они уходят не далее 1 м от входа. Но поскольку гнездо диффузное и входы удалены друг от друга, кормовые участки отдельных секций смыкаются и сравнительно небольшая (видимо, не более 1—5 тыс.) семья эксплуатирует обширную территорию. Общение между отдельными секциями происходит по горизонтальным галереям, расположенным на значительной глубине и поэтому не разрушающимся в конце весны и летом, когда верхние слои почвы на открытых местах совершенно пересыхают и перевиваются ветром. Имеется у M. barbatulum и система поверхностных тоннелей, которая функционирует весной, когда почва бывает влажной.
Сходную функцию выполняет и система тоннелей Monomorium kusnezovi и ![]() M. gracillimum, встречающаяся также у многих муравьев гумидной зоны. Общая площадь территории, занятой тоннелями у M. kusnezovi, порой превышает 10 м². По тоннелям муравьи могут, не появляясь на поверхности, выходить в разных точках кормового участка. Как показал А. А. Захаров (1972), у M. kusnezovi с тоннелями связаны довольно постоянные группы фуражиров, однако при появлении обильного источника пищи вблизи одного из входов происходит перегруппировка и пассивные фуражиры переходят из одного тоннеля в другой. Но в условиях пустыни диффузное гнездо M. barbatulum имеет явные преимущества. Во-первых, у M. kusnezovi пища, доставленная во вход, должна быть затем перенесена в гнездо на большое расстояние, а у M. barbatulum она частично используется в данной секции, так как в каждой секции имеется некоторое количество личинок. И, во-вторых, поверхностные тоннели на открытых местах в рыхлой сухой почве постоянно разрушаются. Поэтому развитая система тоннелей имеется у M. kusnezovi, обитающего во влажной почве солончаков (реже в подстилке саксаулов на краю оазиса) и
M. gracillimum, встречающаяся также у многих муравьев гумидной зоны. Общая площадь территории, занятой тоннелями у M. kusnezovi, порой превышает 10 м². По тоннелям муравьи могут, не появляясь на поверхности, выходить в разных точках кормового участка. Как показал А. А. Захаров (1972), у M. kusnezovi с тоннелями связаны довольно постоянные группы фуражиров, однако при появлении обильного источника пищи вблизи одного из входов происходит перегруппировка и пассивные фуражиры переходят из одного тоннеля в другой. Но в условиях пустыни диффузное гнездо M. barbatulum имеет явные преимущества. Во-первых, у M. kusnezovi пища, доставленная во вход, должна быть затем перенесена в гнездо на большое расстояние, а у M. barbatulum она частично используется в данной секции, так как в каждой секции имеется некоторое количество личинок. И, во-вторых, поверхностные тоннели на открытых местах в рыхлой сухой почве постоянно разрушаются. Поэтому развитая система тоннелей имеется у M. kusnezovi, обитающего во влажной почве солончаков (реже в подстилке саксаулов на краю оазиса) и ![]() M. gracillimum, который в Каракумах тесно связан с подстилкой под саксаулами, а в Каршинской степи встречается в глинистой пустыне или в подстилке в тугаях. Таким образом, гнезда M. barbatulum, как и гнезда Messor variabilis, представляют собой крайний пример специализации гнездостроения, связанной с обитанием в пустынях.
M. gracillimum, который в Каракумах тесно связан с подстилкой под саксаулами, а в Каршинской степи встречается в глинистой пустыне или в подстилке в тугаях. Таким образом, гнезда M. barbatulum, как и гнезда Messor variabilis, представляют собой крайний пример специализации гнездостроения, связанной с обитанием в пустынях.
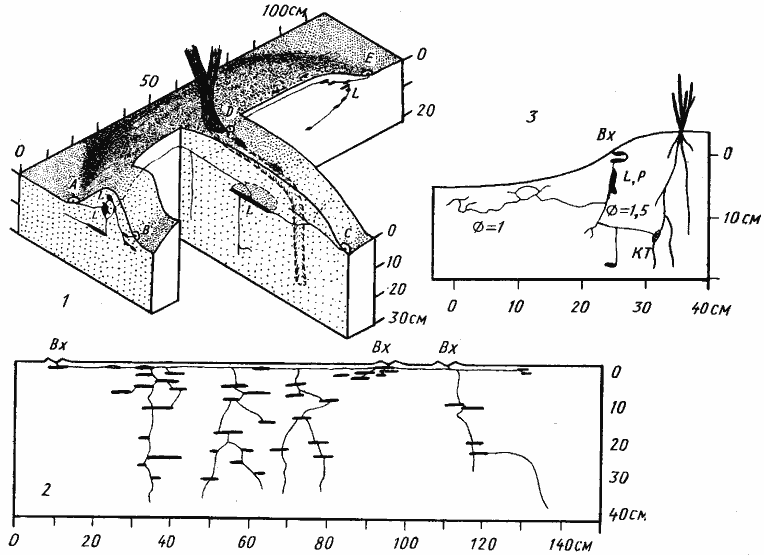
Рис. 65. Гнезда Tetramorium
1 — диффузное гнездо T. schneideri с тремя секциями (входы АВ, CD и Е), Акыбай, № 71-07, 1 апреля 1971 г., в семье около 500 рабочих; 2 — компактное гнездо того же вида с четырьмя линейно расположенными секциями и тремя входами, Акыбай, № 71-20, 24 апреля 1971 г., 2300 рабочих; 3 — небольшое гнездо T. inerme, Акыбай, № 71-09, 2 апреля 1971 г.
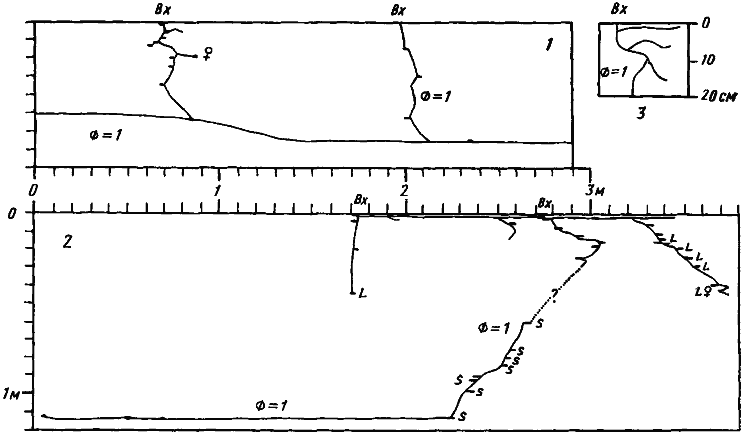
Рис. 68. Гнезда Monomorium barbatulum
1 — фрагмент гнезда № 74-19, Карши, 25 мая 1974 г., лессовидный суглинок, до 30 см сухой, глубже — влажный: 2 — гнездо № 71-28, Акыбай, 6 мая 1971 г., песок, влажный сверху и сухой на глубине более 50 см; 3 — система тонких поверхностных галерей в сухом песке гнезда № 71-06, Акыбай. 29 марта 1971
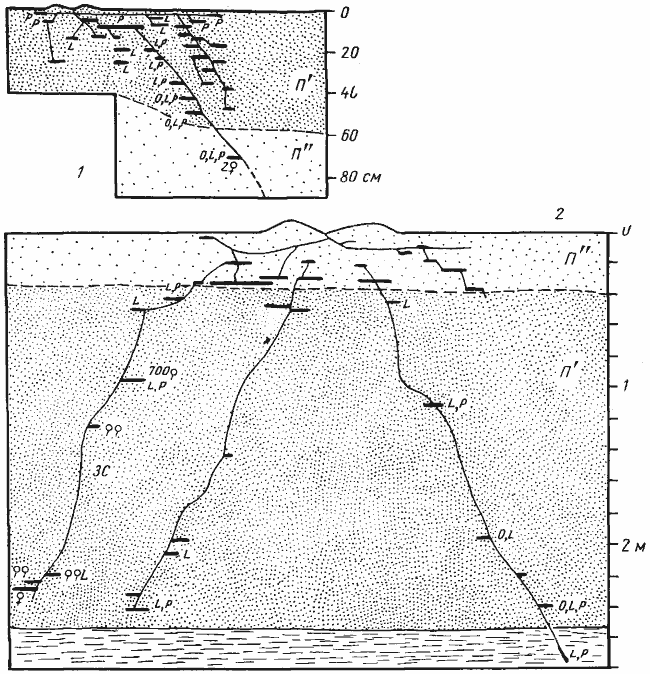
Рис. 60. Односекционные гнезда Cataglyphis setipes
(Акыбай)
1 — 15 мая 1971 г.; ствол засыпан и вся жизнь семьи была сосредоточена в верхнем влажном горизонте почвы; 2 — 1 сентября 1971 г.; ЗС — ствол, камеры которого были забиты рабочими, очевидно уже подготовленными к зимовке
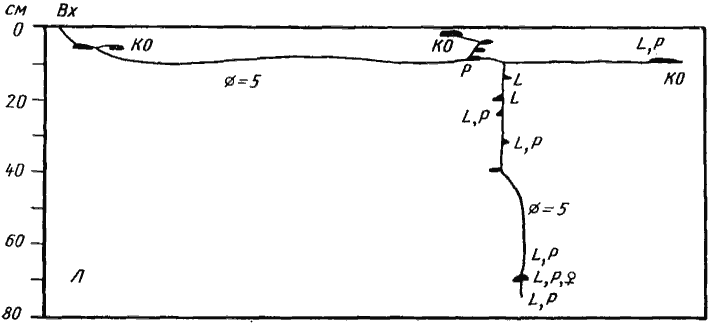
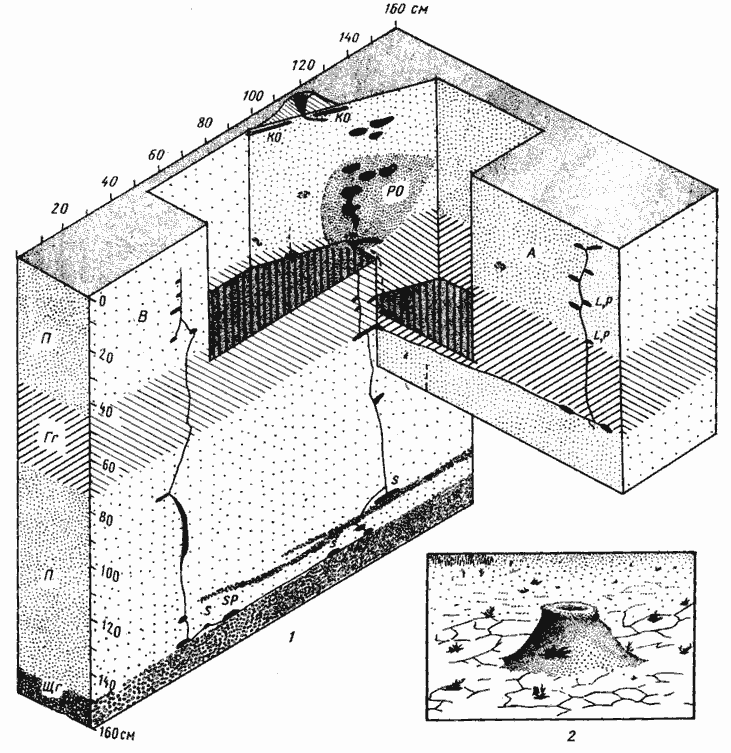
Рис. 63. Гнезда Messor variabilis
1 — Дарья-сай, № 65-145, 27-29 мая 1965 г.; пунктиром обозначены заброшенные камеры и ходы забитые разложившимися органическими остатками; 2 — вход в гнездо Messor variabilis (на такыре, Акыбай)
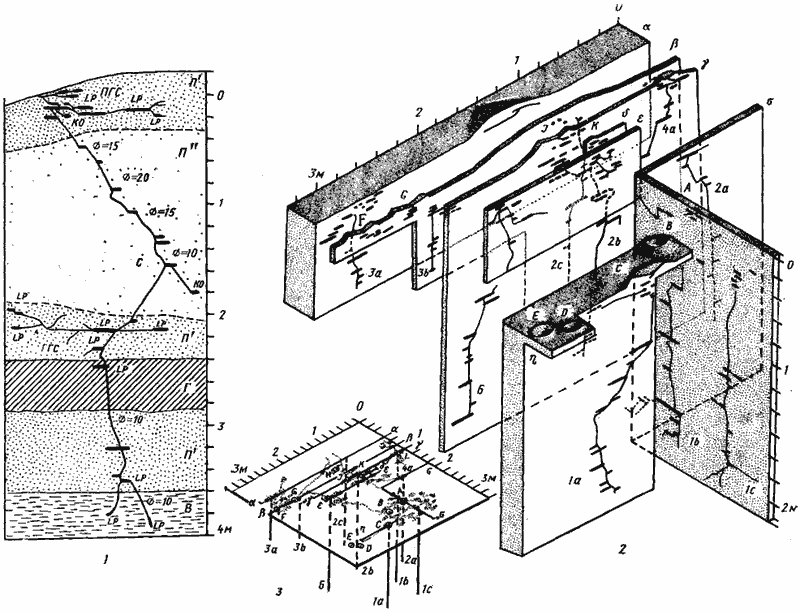
Рис. 67. Два гнезда Camponotus xerxes
1 — Акыбай, № 71-19, 21—22 апреля 1971 г., молодая семья с 859 рабочими; 2, 3 — Акыбай, № 71-11, 8—17 апреля 1971 г. Гнездо с населением около 6 тыс. рабочих, имевшее 6 секций, 11 входов и 10 стволов в пяти раскопанных секциях (секция 5 с входами D и Е не раскапывалась): 2 — изображены те срезы через гнездо, на которых обнажались стволы. Глубина стволов 1а и 1с была соответственно 220 и 240 см. Поскольку при раскопке этих стволов были вскрыты жилы грунтовой воды, вода в яме поднялась вначале до уровня 200 см, а затем — 180 см и поэтому остальные стволы вскрыть до конца не удалось; 3 — это же гнездо изображено схематически. На схеме показано взаимное расположение разрезов, поверхностная горизонтальная система (точками) и стволы гнезда (вертикальными линиями). Разрезы обозначены буквами греческого алфавита, входы в гнездо — заглавными латинскими буквами и стволы — буквенно-цифровым индексом, где цифра означает номер секции. Секция 6 не имела самостоятельного входа и в ней находились самка и основная масса расплода
специальное приспособление для переноски сухой почвы, распространенн у пустынных муравьев, образован длинными отстоящими волосками (аммохетами), часто имеющими форму латинской буквы J, расположен на нижней поверхности головы.
наука изучающая муравьев, часть энтомологии. Мирмекологи - ученые изучающие муравьев.
отряд насекомых. Длина 4 мм 9,5 см. Ок. 3600 видов, преимущественно в тропиках и субтропиках. У ряда видов крылья укорочены или отсутствуют. Некоторые (прусак, черный таракан и др.) обитают в жилище человека. Реликтовый лесной таракан, обитающий на юге Приморья, охраняется.
форма поликалии, при которой семья рассредоточена по готовым камерам и небольшим гнездам, не имеющим постоянного состава. Функциональной дифференциации таких гнезд и камер нет.
необратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.
живут колониями (пчелы, муравьи, термиты, некоторые осы). В состав колонии, напр. у термитов, входят половые особи (самцы и матки обеспечивают размножение), солдаты (защиту) и рабочие (собирание корма, строительство гнезда и т. д.). Солдаты и рабочие бесплодны; половые особи и солдаты сами не способны питаться; таким образом, члены колонии полностью зависят друг от друга. Разделение функций и сложные взаимодействия разных особей в колониях общественных насекомых обеспечиваются рядом особенностей в их строении и поведении; последнее регулируется с помощью выделяемых ими химических веществ (феромонов).
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
выполнение муравьем в течение длительного времени определенных функций в семье. Если функция связана только с возрастом - возрастной полиэтизм; если функция выполняется постоянно - постоянный, или «кастовый», полиэтизм.
обязательное существование только сообществами (семьями, общинами).
Старые названия: Acantholepis semenovi, Acantholepis transcaspiensis
(гр. topos - место) - естественное жизненное пространство определенного биоценоза, совместно с которым он образует экосистему.
Всего 60 видов.
Всего 65 видов.
отряд общественных насекомых. Общины, разделенные на касты, состоят из крылатых и бескрылых особей. Строят подземные и наземные (до 15 м высотой) гнезда (термитники). Ок. 2600 видов, главным образом в тропиках; в России 2 вида: один в районе Сочи, второй, вероятно, завезенный из Японии или Китая, во Владивостоке. Многоядны. Разрушают древесину, портят бумагу, кожу, сельскохозяйственные продукты и др. Термиты, обитающие в почве, полезны как почвообразователи.
гигрофил
ксеробионт
Всего 106 видов.
Всего 965 видов.
организм, предпочитающий влажные местообитания - любящий влажность.
Всего 4 вида.
Старое название: Cataglyphis cinnamomea
гнездо, состоящее из повторяющихся стереотипных блоков (секций), каждый из которых включает все видоспецифичные элементы (ствол, камеры и т. д.)
 Myrmica rubra (Linnaeus, 1758) — рыжая мирмика
Myrmica rubra (Linnaeus, 1758) — рыжая мирмика Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant
Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant Formica cunicularia Latreille, 1798 — прыткий степной муравей
Formica cunicularia Latreille, 1798 — прыткий степной муравей(rp. ontos - сущее, существо) - индивидуальное развитие организма с момента зарождения до смерти.
 Lasius flavus (Fabricius, 1782) — желтый луговой (садовый, земляной) муравей
Lasius flavus (Fabricius, 1782) — желтый луговой (садовый, земляной) муравейорганизационно оформленная мультифункциональная часть семьи (подсистема), имеющая постоянный состав и способная перейти к самостоятельному существованию.
участок, используемый муравьями для добычи пищи.
организм, питающийся семенами растений.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
галобионт
Subspecies: Monomorium indicum kusnezowi Santschi, 1928
 Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушитель
Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушительСтарое название: Monomorium gracillimum
Всего 415 видов
ксеробионт






