






Поведение рабочих особей при постройке гнезд состоит из серий стереотипных действий, которые весьма сходны у представителей одного рода, а часто даже у далеких групп муравьев (Sudd, 1969; Длусский, 1972а). В этом отношении поведение муравьев при рытье ходов напоминает хорошо изученное поведение таких примитивных животных, как личинки ручейников. Их действия можно разбить на стереотипные этапы, причем результат одного этапа служит стимулом для начала следующего (Баранов и др., 1975; Позин, 1972). Правда, сходство это не совсем полное: у муравьев эти элементарные поведенческие блоки относительно изолированы и могут следовать друг за другом в различной последовательности. Так, например, если личинка ручейника отгрызает кусок листа, она скорее всего вслед за этим будет прикреплять этот кусок к домику. У муравьев же один рабочий может часами отгребать почву и лишь в заключение вынести на поверхность ее комок, другой в то же время будет только выносить уже разрыхленную почву и может вообще не копать, третий — в основном разбрасывать вынесенную почву на поверхности. При этом одна и та же особь в один день наблюдений может заниматься преимущественно рытьем, а в другой — выносом почвы на поверхность. Именно поэтому мы предпочитаем термин «блок поведенческих актов», а не «этапы поведения». В пределах такого блока, как и в пределах этапа, поведение, как правило, бывает стереотипным.
Для сравнительного изучения поведения муравьев при прокладке ходов в почве в 1965 г. мы провели серию наблюдений над четырьмя видами Cataglyphis (![]() C. aenescens, C. foreli, C. pallida и C. cinnamomea), Messor aralocaspius и Camponotus turcestanicus на Кызылкумской пустынной станции. 20 рабочих особей помещались в стеклянный узкостенный садок размером 200x300x10 мм, заполненный увлажненной и уплотненной супесчаной почвой. Для выравнивания условий для всех опытов почва бралась из одного, предварительно тщательно перемешанного образца, взятого с глубины 20—40 см. Осенью того же года аналогичные наблюдения были сделаны во Владимирской области над Formica pratensis,
C. aenescens, C. foreli, C. pallida и C. cinnamomea), Messor aralocaspius и Camponotus turcestanicus на Кызылкумской пустынной станции. 20 рабочих особей помещались в стеклянный узкостенный садок размером 200x300x10 мм, заполненный увлажненной и уплотненной супесчаной почвой. Для выравнивания условий для всех опытов почва бралась из одного, предварительно тщательно перемешанного образца, взятого с глубины 20—40 см. Осенью того же года аналогичные наблюдения были сделаны во Владимирской области над Formica pratensis, ![]() Lasius niger,
Lasius niger, ![]() L. flavus,
L. flavus, ![]() Tetramorium caespitum и Leptothorax unifasciatus, но для последних трех видов взяты садки меньшего размера. Почва в этих опытах не выравнивалась. В 1978 г. в лаборатории проводились наблюдения над Alloformica aberrans и
Tetramorium caespitum и Leptothorax unifasciatus, но для последних трех видов взяты садки меньшего размера. Почва в этих опытах не выравнивалась. В 1978 г. в лаборатории проводились наблюдения над Alloformica aberrans и ![]() Cataglyphis aenescens, привезенными из Варганзы, и
Cataglyphis aenescens, привезенными из Варганзы, и ![]() Formica cunicularia, собранными в Московской области. Для опытов с этими тремя видами почва (легкая супесь) также бралась из одного образца. Во всех опытах наблюдения проводились при помощи бинокулярного микроскопа или семикратной лупы. Одновременно с наблюдениями делались зарисовки и фотографии муравьев.
Formica cunicularia, собранными в Московской области. Для опытов с этими тремя видами почва (легкая супесь) также бралась из одного образца. Во всех опытах наблюдения проводились при помощи бинокулярного микроскопа или семикратной лупы. Одновременно с наблюдениями делались зарисовки и фотографии муравьев.
При прокладке ходов в почве у муравьев проявляются следующие блоки поведенческих актов:
1. Рытье (рис:50/1/; рис:51/1/). При собственно рытье почвы ноги муравьев фиксированы и нижнечелюстные щупики прижаты. Все движения совершаются головой и корпусом. Чаще всего при движении к стенке хода муравей одновременно широко раздвигает жвалы, а затем одним движением сжимает их и подается всем корпусом назад, в конце движения снова открывает жвалы и отбрасывает отгрызенный комок под себя. В том случае, если грунт очень плотный (например, при попытках прорваться через стеклянную стенку садка), муравей подается с открытыми жвалами вперед, а затем, прижавшись челюстями к субстрату, начинает быстро открывать и закрывать их. При рытье рыхлой почвы муравей отбрасывает комки почвы под себя передним краем головы, держа жвалы сомкнутыми. Весьма возможно, что этот поведенческий блок на самом деле состоит из нескольких этапов — движения вперед, отгрызания грунта и отбрасывания комка почвы на себя. Тогда действия муравья на очень плотном грунте объясняются удлинением второго этапа. Но детальный анализ без применения скоростной киносъемки затруднен /Провести киносъемку мы не смогли по двум причинам: во-первых, стекло садка всегда сильно загрязнено изнутри и, во-вторых, при сильном освещении, которое требуется для скоростной съемки, муравьи перестают работать./, так как у Formica, Camponotus, Cataglyphis, Lasius и Alloformica эти действия происходят очень быстро и выглядят как одно движение. У Messor aralocaspius поведение при рытье внешне несколько отличается, хотя в принципе остается тем же. Муравей вначале медленно вгрызается в стенку хода, а затем делает резкое движение всем корпусом назад, отрывает небольшой комок, относит его в сторону в пределах хода и снова возвращается на место. Мелкие муравьи (Lasius, Tetramorium, Leptothorax) обычно просто вытаскивают из стенки хода несколько песчинок и относят в сторону.
2. Отгребание (рис:50/2/; рис:51/2/). Отделенные от стенки хода комочки почвы отгребаются назад по уже прорытому ходу. При этом используется два различных приема:
а) опустив голову со сложенными жвалами, муравей делает движения головой под себя и одновременно толчками продвигается назад;
б) муравей подгребает почву под себя передними ногами и одновременно толчками движется назад.
Иногда эти два приема используются вместо рытья при прокладке ходов в очень рыхлой почве. Особенно часто второй прием наблюдается у ![]() Tetramorium caespitum.
Tetramorium caespitum.
3. Формовка комка почвы (рис:50/3/; рис:51/3/). Виды, не имеющие псаммофора, сгребают в комок рыхлую почву с пола хода головой (движениями под себя) и передними ногами (с боков к себе, используя широкую нижнюю поверхность последних члеников лапок). Мелкие муравьи часто подбирают с пола комок слипшихся песчинок и выносят наверх. У видов, имеющих псаммофор, упаковка в него почвы происходит по-разному в разных родах. Подробно упаковка псаммофора описана выше.
4. Транспортировка комка почвы (рис:50/4/; рис:51/4/).
5. Освобождение от комка почвы. Большинство муравьев просто разводят жвалы и роняют принесенный комок. Действия видов, имеющих псаммофор, сложнее. Рабочие Cataglyphis широко раскрывают жвалы и одновременно отгибают назад нижнечелюстные щупики и делают короткий толчок головой или всем телом. Рабочие Messor «вычесывают» песчинки из бороды, проводя передними лапками вперед.
6. Трамбовка. Брошенный (или положенный) комок почвы муравей быстро «притопывает» короткими ударами ног; работают при этом последние членики лапок. Чаще всего таким образом рабочие разбивают комки почвы, вынесенные на поверхность, но Lasius и Tetramorium таким образом вминают почву в стенки камер и ходов, уплотняя их. Интересно, что у A. aberrans, у которого в 60% случаев рабочие «растаптывают» комки почвы, вынесенные на поверхность, эту операцию невозможно временами отличить от короткой чистки передних лапок. Иногда рабочие, бросив комок, отходят в сторону, а затем чистятся. Но иногда чистка следует непосредственно за освобождением челюстей (когда голова еще наклонена к земле) и, проводя лапками через максиллы, муравей одновременно толкает комок почвы. В некоторых же случаях рабочие этого вида явно трамбуют комок и голова при этом приподнята.
7. Заталкивание комка. Иногда у Cataglyphis cinnamomea рабочие заполняют почвой щели между камнями. Муравей кладет комок возле щели и заталкивает закрытыми жвалами резкими движениями головы.
8. Отбрасывание почвы (рис:50/5/). Муравей, вынесший комок, поворачивается головой к гнезду и начинает, продвигаясь назад, быстро отбрасывать назад почву передними ногами. Из-под задних ног при этом вылетает струйка песка. Такое поведение очень характерно для многих роющих ос. Среди муравьев мы наблюдали его только у представителей трибы Formicini, причем в ярко выраженной форме только у пустынных Cataglyphis. Такое же поведение описывается у Ocymyrmex пустынь Южной Африки (Sudd, 1967).
9. Чистка (рис:50/6-10/; рис:51/5,6/). Время от времени муравьи прерывают работу и чистятся. Чаще всего чистка происходит после того, как рабочий выносит и бросает комок почвы. Муравьи тщательно чистят шпорами передних голеней усики и лапки средних и задних ног. Передние ноги чаще всего чистят пропуская их через максиллу.
10. Возвращение в гнездо.
11. Вылизывание ходов. Наблюдается не у всех видов. Обычно этим занимаются особые рабочие, которые (во всяком случае в период наблюдений) строительством не занимаются. Прижав нижнюю губу к стенке, муравей совершает очень быстрые движения язычком. На стекле при этом остаются капли жидкости. Следует отметить, что хотя язычок муравья движется по стеклу, мельчайшие песчинки, прилипшие к стеклу, даже не сдвигаются с места. Так что движения в этом случае совсем не такие, как у языка позвоночных животных. Скорее всего это очень частые «пришлепывания», а не «лизание» языком.
Этим списком практически исчерпываются все действия, которые можно наблюдать у изученных муравьев при прокладке ходов в почве. Но реальный поведенческий репертуар каждого отдельного вида еще беднее. Сравнение разных видов (табл.21) показывает, что наибольшие различия наблюдаются у представителей разных родов.
Так, для Cataglyphis характерно отбрасывание почвы и своеобразное поведение при упаковке псаммофора. Мы ни разу не наблюдали, чтобы эти муравьи вылизывали стенки ходов. Сходно поведение Alloformica и Formica, но у последних отмечалось вылизывание стенок ходов. Camponotus turcestanicus никогда не трамбует и не отгребает почвы ногами. Напротив, Messor aralocaspius трамбует почти каждый комок почвы, вынесенный из гнезда. Наиболее простое поведение наблюдалось у мелких муравьев — Lasius, Tetramorium и Leptothorax.
В пределах рода разные виды имеют, как правило, одинаковый набор поведенческих блоков. Различия заключаются в том, что каждый вид чаще использует одни приемы, чем другие. Так, например, у Cataglyphis при прокладке ходов в одинаковой почве (опыты 1965 г.) внутри гнезда поведение всех четырех видов не отличалось, но на поверхности поведение было различным у разных видов. Так, ![]() C. aenescens, обитающий на влажных почвах солончаков и оазисов, преимущественно рыхлых, сразу же после освобождения псаммофора или предварительно почистившись обязательно отбрасывал почву. То же характерно и для C. pallida — обитателя песчаных пустынь, но у этого вида отбрасывание занимает гораздо больше времени, чем у всех остальных муравьев. В естественных условиях у входов в гнездо этого вида постоянно находится несколько рабочих, отбрасывающих песок от входа. Если такие рабочие, испугавшись, скрываются в гнезде, вход начинает в сухую жаркую погоду быстро заплывать оползающим сухим песком. Рабочие C. foreli в условиях данного опыта отбрасывали почву лишь при выкапывании засыпанных (при заселении садка муравьями) личинок. C. cinnamomea вообще никогда не отбрасывали почвы, но зато заталкивали комки почвы в различные трещины и щели.
C. aenescens, обитающий на влажных почвах солончаков и оазисов, преимущественно рыхлых, сразу же после освобождения псаммофора или предварительно почистившись обязательно отбрасывал почву. То же характерно и для C. pallida — обитателя песчаных пустынь, но у этого вида отбрасывание занимает гораздо больше времени, чем у всех остальных муравьев. В естественных условиях у входов в гнездо этого вида постоянно находится несколько рабочих, отбрасывающих песок от входа. Если такие рабочие, испугавшись, скрываются в гнезде, вход начинает в сухую жаркую погоду быстро заплывать оползающим сухим песком. Рабочие C. foreli в условиях данного опыта отбрасывали почву лишь при выкапывании засыпанных (при заселении садка муравьями) личинок. C. cinnamomea вообще никогда не отбрасывали почвы, но зато заталкивали комки почвы в различные трещины и щели.
Следует еще раз подчеркнуть, что описанные наблюдения сделаны при прокладке ходов в одной и той же почве. Поведение одного вида при работе в разных почвах будет различным. Так, например, при помещении ![]() C. aenescens в почву другого состава (песок не эолового, а наносного происхождения, т.е. более крупнозернистый) в опытах 1978 г. рабочие этого муравья отбрасывали почву только в 20—30% выходов из гнезда, а не 90—100%, как в описанном выше опыте. В естественных условиях в Каррыкуле C. foreli в гнездах на барханах систематически отбрасывает почву у гнезд, а на такыровидных почвах никогда этого не делает. Но различия между видами сохраняются и при переходе на другой тип почвы. Так, в опытах 1978 г. A. aberrans, помещенные в ту же почву, что и
C. aenescens в почву другого состава (песок не эолового, а наносного происхождения, т.е. более крупнозернистый) в опытах 1978 г. рабочие этого муравья отбрасывали почву только в 20—30% выходов из гнезда, а не 90—100%, как в описанном выше опыте. В естественных условиях в Каррыкуле C. foreli в гнездах на барханах систематически отбрасывает почву у гнезд, а на такыровидных почвах никогда этого не делает. Но различия между видами сохраняются и при переходе на другой тип почвы. Так, в опытах 1978 г. A. aberrans, помещенные в ту же почву, что и ![]() C. aenescens, отбрасывали почву только в 2% выходов, но зато в 59% выходов трамбовали вынесенные комки, что
C. aenescens, отбрасывали почву только в 2% выходов, но зато в 59% выходов трамбовали вынесенные комки, что ![]() C. aenescens делает очень редко (менее 1%). Отличаются и пределы условий, в которых может работать тот или иной вид. Так, в сухом песке A. aberrans,
C. aenescens делает очень редко (менее 1%). Отличаются и пределы условий, в которых может работать тот или иной вид. Так, в сухом песке A. aberrans, ![]() F. cunicularia и C. cinnamomea просто не способны рыть ходы, а C. pallida прекрасно справляется с постройкой в этой ситуации.
F. cunicularia и C. cinnamomea просто не способны рыть ходы, а C. pallida прекрасно справляется с постройкой в этой ситуации.
Различия наблюдаются и у особей одной семьи, особенно в разных ситуациях. Так, например, у ![]() C. aenescens, если в прокладке хода участвуют один-три рабочих, они выполняют весь поведенческий репертуар и последовательность поведенческих блоков будет
C. aenescens, если в прокладке хода участвуют один-три рабочих, они выполняют весь поведенческий репертуар и последовательность поведенческих блоков будет
(1,1...1→3→4→5→8→9→10)1..n или (1,1...1→3→4→5→9→8→10)1..n
Если же в ходе работает большее число особей, то происходит разделение функций и наблюдается два типа последовательности:
(1) (1,1...1→2a...2a→9)1..n
и (2) (3→4→5→8→9→10)1..n или (3→4→5→9→8→10)1..n
иногда к ним добавляется третья группа особей
(3) (8,8...8→4*→9)1..n
У M. aralocaspius при прокладке ходов немногими рабочими последовательность действий обычно бывает
(9→(1↔4*)...(1↔4*)→3→4→5→6→10)1..n
а при работе группы особей —
(1) ((1↔4*)...(1↔4*)→9)1..n или ((1↔4*)→9→(1→4*→9)1..n)1..n
и (2) (9→9→4→6→10)1..n
/* Транспортировка частиц почвы в жвалах на небольшое (несколько сантиметров) расстояние./
Как видно из табл.21, поведение при прокладке ходов у муравьев, обитающих в пустыне, мало отличается от поведения видов гумидных областей. Разница сводится в основном к действиям по упаковке и очистке псаммофора и усилению (по сравнению с другими представителями трибы Formicini) эффективности отбрасывания почвы у Cataglyphis. Отсюда можно сделать вывод, что поведенческие стереотипы, сложившиеся у муравьев в гумидных областях, оказались в основном пригодными для постройки гнезд в условиях пустыни. Менялся главным образом характер реакции муравьев на тип (точнее — механический состав) почвы, выражающийся в преобладании различных стереотипов поведения у разных видов при прокладке ходов в одних и тех же условиях.
Не следует, однако, думать, что вся гнездостроительная деятельность муравьев может быть сведена к набору стереотипных блоков. Дж. Садд (Sudd, 1975), изучавший прокладку ходов у Formica lemani и ![]() Lasius niger, построил машинную программу, моделирующую постройку гнезда муравьями. Полученные на ЭВМ результаты хорошо описывали прокладку хода у F. lemani, но не предсказывали ни ветвления ходов, ни постройки камер. И пока еще мы очень далеки от того, чтобы понять, каким образом в результате этих действий (в общем очень простых) появляется сложное и совершенное гнездо, имеющее, с одной стороны, специфические особенности у каждого вида, а с другой — приспособленное к конкретному местообитанию.
Lasius niger, построил машинную программу, моделирующую постройку гнезда муравьями. Полученные на ЭВМ результаты хорошо описывали прокладку хода у F. lemani, но не предсказывали ни ветвления ходов, ни постройки камер. И пока еще мы очень далеки от того, чтобы понять, каким образом в результате этих действий (в общем очень простых) появляется сложное и совершенное гнездо, имеющее, с одной стороны, специфические особенности у каждого вида, а с другой — приспособленное к конкретному местообитанию.
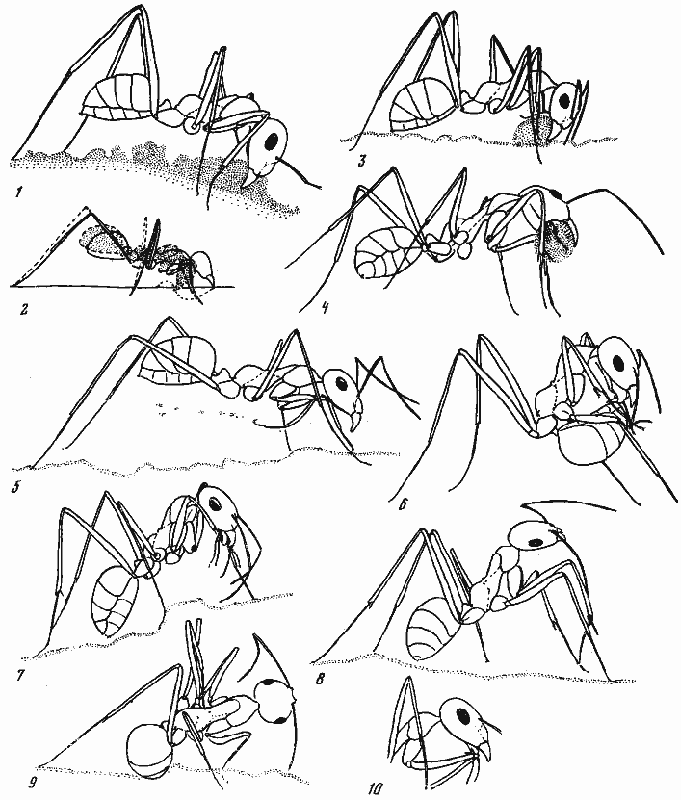
Рис. 50. Поведение Cataglyphis foreli при прокладке ходов
(прорисовки по фотографиям)
1 — отгребание почвы; 2 — движения муравья при отгребании почвы; 3 — формовка комка; 4 — транспортировка почвы; 5 — отбрасывание почвы передними ногами; 6—10 — различные приемы чистки: 6 — чистка конца брюшка, 7 — чистка лапки при помощи максилл, 8 — чистка жгутика усика чистильным аппаратом, образованным основанием первого членика передней лапки и модифицированной шпорой передней голени, 9 — чистка лапки средней ноги тем же чистильным аппаратом, 10 — чистка коленного сочленения
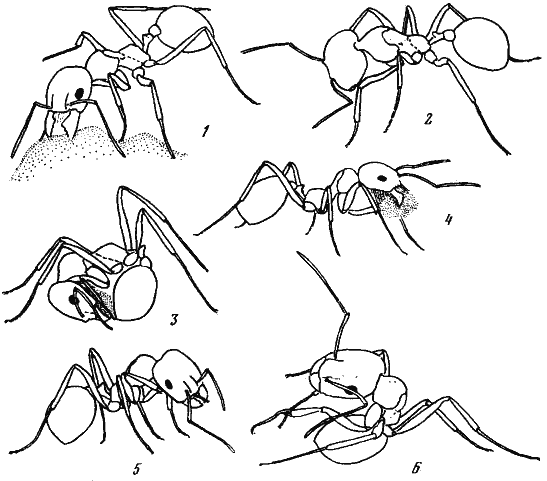
Рис. 51. Поведение Messor aralocaspius при прокладке ходов
(прорисовки по фотографиям)
1 — отгрызание твердой почвы; 2 — отгребание почвы; 3 — формовка комка; 4 — транспортировка; 5 — чистка передней лапки максиллой; 6 — чистка средней лапки чистильным аппаратом
Всего 65 видов.
 Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунок
Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунокгигрофил, галобионт
Старое название: Cataglyphis flavigastra
ксеробионт
Старое название: Cataglyphis cinnamomea
ксеробионт
 Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant
Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant Lasius flavus (Fabricius, 1782) — желтый луговой (садовый, земляной) муравей
Lasius flavus (Fabricius, 1782) — желтый луговой (садовый, земляной) муравей Tetramorium caespitum (Linnaeus, 1758) — дерновый муравей, pavement ant
Tetramorium caespitum (Linnaeus, 1758) — дерновый муравей, pavement antСтарое название: T. indocile
Старое название: Leptothorax unifasciatus
 Formica cunicularia Latreille, 1798 — прыткий степной муравей
Formica cunicularia Latreille, 1798 — прыткий степной муравей(лат. mandibule — челюсть, от mando — жую, грызу), первая пара челюстей у ракообразных, многоножек и насекомых. Синоним: жвала.
Всего 965 видов.
Всего 100 видов.
Всего 4 вида.
Всего 415 видов
Всего 19 видов.
специальное приспособление для переноски сухой почвы, распространенн у пустынных муравьев, образован длинными отстоящими волосками (аммохетами), часто имеющими форму латинской буквы J, расположен на нижней поверхности головы.
Всего 106 видов.
сборное название насекомых подотряда жалящих перепончатокрылых (кроме пчел и муравьев). Длина до 40 мм (шершни). Ок. 23 тыс. видов. Распространены широко. Опылители растений. Личинки некоторых ос паразиты других насекомых.
восприятие муравьями в качестве сигнала характерных движений других особей одной общины («язык поз»).
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).






