






Способность создавать запасы пищи в большей или меньшей степени присуща всем муравьям среднеазиатских пустынь. Исключение представляют виды, обитающие во влажных биотопах (или микростациях) и имеющие постоянные источники пищи в течение сезона активной жизни (Solenopsis, Leptothorax, Tapinoma). Хотя настоящие медовые муравьи здесь отсутствуют (исключение — Proformica epinotalis северных пустынь), все пустынные![]() Formicinae, как уже говорилось, могут накапливать в зобиках значительные количества жидкой пищи. Хорошо известен присущий большинству пустынных
Formicinae, как уже говорилось, могут накапливать в зобиках значительные количества жидкой пищи. Хорошо известен присущий большинству пустынных![]() Myrmicinae инстинкт запасания семян. У Cataglyphis foreli и Crematogaster subdentata мы также обнаружили способность делать запасы из высушенных насекомых (Длусский, 1971, 1975а).
Myrmicinae инстинкт запасания семян. У Cataglyphis foreli и Crematogaster subdentata мы также обнаружили способность делать запасы из высушенных насекомых (Длусский, 1971, 1975а).
Предпосылкой для возникновения специализированного запасания корма у муравьев является способность большинства видов гумидных областей делать кратковременные запасы при изобилии пищи на кормовом участке. Такая способность была обнаружена нами впервые у ![]() Myrmica rubra (Длусский, 19676; Dlussky, Kupianskaya, 1972), а затем мы наблюдали ее у M. ussuriensis, M. schencki, M. scabrinodis, Aphaenogaster subterranea,
Myrmica rubra (Длусский, 19676; Dlussky, Kupianskaya, 1972), а затем мы наблюдали ее у M. ussuriensis, M. schencki, M. scabrinodis, Aphaenogaster subterranea, ![]() Lasius niger, Camponotus herculeanus, C. aethiops и других видов, с которыми нам приходилось работать в лаборатории. Если давать муравьям избыточное количество белковой пищи, то количество корма, унесенного за день, сильно флуктуирует. В два соседние дня при стабильных условиях температуры, влажности и освещенности эти количества могут различаться в 10 раз. Обычно муравьи набирают больше корма, чем могут съесть их личинки, и складывают в камерах гнезда, а за новым кормом перестают выходить. Чаще всего часть этого запасенного корма портится и выбрасывается муравьями из гнезда. После того как большая часть запасенной пищи используется, начинается новый цикл. У
Lasius niger, Camponotus herculeanus, C. aethiops и других видов, с которыми нам приходилось работать в лаборатории. Если давать муравьям избыточное количество белковой пищи, то количество корма, унесенного за день, сильно флуктуирует. В два соседние дня при стабильных условиях температуры, влажности и освещенности эти количества могут различаться в 10 раз. Обычно муравьи набирают больше корма, чем могут съесть их личинки, и складывают в камерах гнезда, а за новым кормом перестают выходить. Чаще всего часть этого запасенного корма портится и выбрасывается муравьями из гнезда. После того как большая часть запасенной пищи используется, начинается новый цикл. У ![]() M. rubra длительность циклов (от пика до пика) составляет от трех до шести дней (рис:43).
M. rubra длительность циклов (от пика до пика) составляет от трех до шести дней (рис:43).
Сходное кратковременное запасание корма при его изобилии, видимо, характерно для большинства животных, имеющих гнезда (в широком смысле этого термина). П. А. Свириденко (1957), собравший и обобщивший большой материал по эволюции инстинкта запасания пищи у позвоночных животных, описывает сходное поведение у некоторых грызунов. Автор также рассматривает такой тип поведения, сочетающийся у одиночных животных со стремлением спрятать пищу от врагов, яркого света или неблагоприятной температуры (по терминологии автора — кинестетическая реакция на корм в сочетании с пассивно-оборонительной реакцией), как базу для возникновения инстинкта запасания.
В условиях гумидного климата эта способность муравьев не может получить достаточного развития и превратиться в инстинкт запасания, так как их обычная добыча (трупы насекомых) не может долго сохраняться во влажной почве. В 1976 г. в лабораторных условиях мы предлагали Tetramorium taurocaucasum куски мяса. В естественных условиях эти муравьи могут делать небольшие запасы семян. Рабочие охотно расчленяли мясо, несли большое количество фрагментов в гнездо и складывали в камерах. Однако вскоре они начали выносить испорченные частицы из гнезда и через два дня потоки приноса и выноса почти уравнялись.
В условиях пустыни в сухой почве высушенные трупы насекомых мотут сохраняться долго. Поэтому здесь у Cataglyphis foreli и Crematogaster subdentata эта способность получает дальнейшее развитие. Запасы пищи в гнездах C. foreli невелики — всего один-два десятка трупов термитов, пустынных мокриц или других крупных беспозвоночных. Обычно этот запас располагается в горизонтальной галерее, проходящей под самой поверхностью почвы. В лабораторном гнезде была сделана проверка, показавшая, что личинки этих муравьев поедают такие высушенные трупы и нормально развиваются.
У Cr. subdentata относительные размеры запасов гораздо больше. Эти муравьи тесно связаны с тлями (в Акыбае — Brachyunguis saxaulica на саксауле) и посещают их для получения пади, но в то же время убивают большое количество тлей и высушивают их в камерах гнезда. Чаще всего такие камеры с запасами делаются в сухих ветках, реже в почве или под камнями. В одной камере может находиться несколько тысяч высушенных тлей. Изредка попадаются другие насекомые — обычно мелкие мухи и личинки цикадок.
Но наиболее ярко способность к запасанию пищи появилась у видов, перешедших к карпофагии (питание семенами). Питание семенами зарегистрировано у многих муравьев гумидной зоны, однако в этих условиях они поедают не сами семена, а различные съедобные придатки семян (эласмосомы). Возникновение таких придатков — результат приспособления растений к расселению муравьями (мирмекохории). В лесах умеренной зоны многие растения нижнего яруса расселяются таким путем (Sernander, 1906). По всей видимости, в условиях недостатка корма некоторые![]() Myrmicinae, не способные запасать пищу в зобу, перешли от питания эласмосомами к поеданию собственно семян. Скорее всего этот переход происходил неоднократно у разных видов в степях и саваннах. Карпофагия, облигатная или факультативная, очень широко распространена среди
Myrmicinae, не способные запасать пищу в зобу, перешли от питания эласмосомами к поеданию собственно семян. Скорее всего этот переход происходил неоднократно у разных видов в степях и саваннах. Карпофагия, облигатная или факультативная, очень широко распространена среди![]() Myrmicinae аридных и субаридных областей.
Myrmicinae аридных и субаридных областей.
Семена представляют собой необычайно удобный материал для длительного хранения. Они не портятся даже в довольно влажных камерах гнезд муравьев-жнецов, находящихся на глубине 2—3 м. Поэтому на базе карпофагии легко возникает, независимо в разных трибах муравьев, инстинкт создания запасов. Однако далеко не все виды, способные поедать семена, могут делать значительные запасы. Так, например, хотя у европейского ![]() Tetramorium caespitum семена составляют значительную часть пищевого рациона (Brian et al., 1967), настоящих запасов в гнездах в Европе эти муравьи обычно не делают. Иногда можно обнаружить лишь некоторое количество семян в камерах с расплодом. Точно так же лишь очень небольшие запасы, которых явно не может хватить на долгое время, в пустынях Средней Азии делают Tetramorium inerme и Pheidole arenarum. У
Tetramorium caespitum семена составляют значительную часть пищевого рациона (Brian et al., 1967), настоящих запасов в гнездах в Европе эти муравьи обычно не делают. Иногда можно обнаружить лишь некоторое количество семян в камерах с расплодом. Точно так же лишь очень небольшие запасы, которых явно не может хватить на долгое время, в пустынях Средней Азии делают Tetramorium inerme и Pheidole arenarum. У ![]() Monomorium gracillimum, M. kusnezovi, Leptothorax semenovi и большинства пустынных видов Tetramorium к концу весны в гнездах накапливаются запасы семян, чаще всего в специальных камерах у самой поверхности почвы. 15—31 мая 1978 г. в окрестностях Варганзы (Узбекистан) мы собрали запасы из пяти гнезд Tetramorium striativentre, которые оказались равными 20, 187, 218, 228 и 330 мг (среднее — 196 мг). Сходная картина наблюдалась и у T. turcmenicum. В трех гнездах этого вида, раскопанных в том же районе, запасы семян составляли 245, 289 и 517 мг.
Monomorium gracillimum, M. kusnezovi, Leptothorax semenovi и большинства пустынных видов Tetramorium к концу весны в гнездах накапливаются запасы семян, чаще всего в специальных камерах у самой поверхности почвы. 15—31 мая 1978 г. в окрестностях Варганзы (Узбекистан) мы собрали запасы из пяти гнезд Tetramorium striativentre, которые оказались равными 20, 187, 218, 228 и 330 мг (среднее — 196 мг). Сходная картина наблюдалась и у T. turcmenicum. В трех гнездах этого вида, раскопанных в том же районе, запасы семян составляли 245, 289 и 517 мг.
Чтобы оценить значение этих цифр, необходимо рассмотреть значение некоторых параметров и индексов, используемых в экологии продуктивности. Под чистой продукцией (NP) понимается суммарный сухой вес рабочих и половых особей, выведенных за определенную единицу времени (обычно за год). Потребление (J) — это суммарный сухой вес пищи за то же время. Стационарная биомасса (В) — средний сухой вес всего населения (для муравьев — практически сухой вес рабочих особей) (Длусский, 1975а). Использование энергетических единиц (калории, джоули) для муравьев нецелесообразно, так как основная масса пищи идет на выкармливание малоподвижных личинок, для которых решающее значение имеет не энергетическая ценность, а качество пиши (количество белка, соотношение аминокислот). Зная величину индекса NP/B, мы можем оценивать продукцию, измеряя биомассу семей. Величина этого индекса у муравьев колеблется от 0,3 до 1,4 и зависит от смертности и доли выведенных крылатых половых особей. Экологическая эффективность роста (NP/J) показывает, сколько единиц пищи необходимо для производства единицы продукции. У хищных беспозвоночных эта величина варьирует в пределах 0,1—0,5, у растительноядных — 0,08—0,27 и у сапрофагов — 0,05—0,08. Все ссылки на величину индексов, приведенные в тексте данного раздела, даны по обзору И. Пентал (Petal, 1978) /В ссылке на работу (Golley, Gentry, 1964) в обзоре И. Пентал имеется опечатка: у Pogonomyrmex badius, по данным этих авторов, NP/J=0,03./.
У T. striativentre численность населения семей 1—5 тыс. особей и биомасса 0,4—2 г. Даже если принять самое высокое значение NP/J, известное для фитофагов (0,27), и самое высокое значение NP/B, полученное для муравьев (1,4), запаса в 200 мг семян достаточно лишь для того, чтобы обеспечить 4—20% потребностей семей. Все эти виды сочетают карпофагию с некрофагией или зоофагией.
Настоящие запасы семян, позволяющие обеспечить питанием семью на весь сухой сезон и выкормить к осени крылатых половых особей, создают только облигатные карпофаги, к которым в пустынях Средней Азии относятся Monomorium barbatulum и все виды Messor. Правда, Messor иногда собирают трупы насекомых и даже ловят мелких или малоподвижных беспозвоночных (тли, поденки, мелкие нимфы цикадок, гусеницы), но роль пищи животного происхождения в их питании ничтожна. У наиболее «хищного» из среднеазиатских муравьев-жнецов M. rufitarsis darianus насекомые и их трупы составляли (по весу) 7,3% (Варганза, объединенные данные нескольких дней наблюдений за несколькими гнездами в конце мая 1978 г.). У M. aralocaspius насекомые в питании практически отсутствуют (менее 0,1% в Акыбае за все время наблюдений); M. denticulatus занимает промежуточное положение между этими видами. Характерная особенность гнезд всех видов облигатных карпофагов — наличие специальных камер для запасания семян, расположенных обычно на значительной глубине.
Хотя запасание семян муравьями-жнецами известно с глубокой древности, количественные данные о размерах запасов и о динамике запасания и расходования пищи крайне скудны. Это связано, несомненно, с трудностью раскопки гнезд, особенно в аридных районах, где гнезда этих муравьев уходят вглубь на несколько метров. Даже в районах с легкой песчаной почвой, где грунтовые воды подходят к поверхности достаточно близко (2—4 м в Акыбае), раскопка одного гнезда занимает несколько дней. Сравнительно недавно появилась первая работа, где обстоятельно разбирается этот вопрос (Lavigne, 1969). Автор ежемесячно в течение года раскапывал по одному гнезду американского муравья-жнеца Pogonomyrmex occidentalis в прерии на высоте около 1500 м в окрестностях Робертона (Вайоминг, США). Правда, выбранная методика не позволила четко проследить динамику запасания пищи, так как взятые в различные сроки семьи сильно отличались размерами (от 691 до 8796 рабочих), однако общие тенденции на материале автора хорошо прослеживаются.
При средней численности семей 4970 особей средние размеры запасов семян составляли 24,5 г. С ноября по май запасы в разных гнездах варьировали от 0 до 23 г, причем в основном состояли из крупных семян Chenopodium. В июле и августе с наступлением влажного сезона размеры запасов явно возрастают: гнездо, раскопанное в июне, имело население 7,5 тыс. особей и запас 33 г, в июле — 5,1 тыс. и 54,3 г и в августе — 5,6 тыс. особей и 66,9 г. При этом в июле и августе основу запаса составляли мелкие семена Capseila и Lepidiutn. В этот же период происходило и развитие расплода (июнь — сентябрь). В октябре и ноябре запасы в раскопанных гнездах были меньше, чем в июле (25,9 и 44,2 г), но сбор семян явно продолжался, о чем свидетельствует появление в камерах с запасами семян не встречавшихся раньше видов растений.
Отдельные сведения имеются и по другим видам Pogonomyrmex. Галли и Джентри (Golley, Gentry, 1964), раскапывая четыре гнезда P. badius, обнаружили в них запасы, равные 16, 73, 130 и 330 г. Исследования, проведенные в пустыне Чиуауа (Whitford, Ettershank, 1975; Whitford, 1978), показали, что у обитающих здесь видов Pogonomyrmex (три вида) особи из всех гнезд работают только один-два месяца за сезон. В остальное время либо открывается только часть гнезд, либо все гнезда бывают закрытыми. Авторы показали, что основным фактором, определяющим сезонную динамику активности фуражировки, является обилие созревших семян на территории.
В 1970—1971 гг. нами была предпринята работа по изучению экологии муравьев-жнецов Туркмении /Сбор материала в значительной мере проводился совместно с К. Сапарлиевым, тогда аспирантом Института зоологии АН ТССР, и частично опубликован (Сапарлиев, 1972; Длусский, 1975а; Длусский, Сапарлыев, 1975)./, одной из задач которой было выяснение динамики сбора пищи. Работа в основном проводилась на двух пробных площадках в Акыбайском лесничестве (Марыйский район Туркмении). Одна из площадок представляла участок с такыровидной почвой среди саксаульника, площадью 734 м², на которой находилось девять гнезд Messor aralocaspius и два гнезда M. intermedius. На участке имелись пятна песчаной почвы (общая их площадь 170 м², или 27% от всей площадки), зарастающие весной эфемерами. Динамика плодоношения эфемеров на этом участке показана на рис:44. Вторая площадка располагалась в 2 км от первой на берегу арыка на засоленном участке с сорной растительностью и редкими кустами — Tamarix. В отличие от площадки №1 здесь наблюдалось осеннее плодоношение растений (некоторые солянки). На площадке было выбрано для наблюдений девять гнезд M. denticulatus. Дважды в месяц определялась суточная активность муравьев, количество фуражировочных экскурсий на гнездо в сутки, процент работающих семей и процент рабочих, возвращающихся в гнездо с семенами.
Характерной особенностью Messor является то, что их гнезда периодически закрываются и муравьи перестают работать на поверхности. В каждом гнезде периоды активности сменяются периодами покоя. Сходные флуктуации обнаружены и у американских муравьев-жнецов в пустыне Чиуауа (Whitford, 1976). От соотношения длительности этих периодов зависит такой показатель, как ежедневное соотношение активных и покоящихся семей. Гнезда могут быть и открытыми, но при этом муравьи работают только на поверхности гнезд и в непосредственной близости от них, а дорог не делают. Обычно в это время муравьи занимаются ремонтом и расширением гнезд. В течение сезона ежедневное соотношение активных и покоящихся семей, средняя интенсивность фуражировки и интенсивность фуражировки активных муравейников сильно меняются.
У M. aralocaspius (рис:45) муравейники пробуждаются в марте. Ежедневно открывается 60—70% гнезд, однако фуражировка еще не начинается. В апреле ежедневно открывалось 15—25% муравейников, но, несмотря на малое количество работающих семей, их активность велика (18,7 тыс. экскурсий на гнездо в сутки в 1970 г. и 6,7 тыс. — в 1971 г.). Довольно высока и эффективность фуражировки — 79,7% фуражиров возвращаются в гнездо с семенами. За день рабочие одной семьи в среднем собирали в 1970 г. 1,49 г семян эфемеров. Часть семей (15% в 1971 г.) продолжает работать на поверхности гнезд и не делает фуражировочных дорог. С мая по август работы на поверхности гнезда практически прекращаются или осуществляются небольшой группой рабочих-строителей во время фуражировки.
Максимум фуражировочной активности отмечался у этого вида в мае 1970 г. В это время фуражировкой занимались ежедневно 80% семей и, хотя активность работающих семей снизилась, среднее количество экскурсий на гнездо возросло по сравнению с апрелем. Увеличилась и эффективность фуражировки: с семенами возвращались 81,4% фуражиров. В среднем рабочие одной семьи приносили в гнезда в этом месяце 2,98 г семян. В последующие месяцы наблюдалось снижение фуражировочной активности. В июне без добычи возвращались в гнездо уже около 50% фуражиров, а в июле и августе — более 70%. За день рабочие одного гнезда приносили в июне 0,15, а в июле — 0,12 г семян. В сентябре фуражировка прекратилась полностью и не возобновлялась до следующей весны. Этот перерыв явно объясняется не погодными условиями или эндогенными факторами, а связан с отсутствием пищи на территории. Когда осенью 1971 г. мы выкладывали на такыре приманки, муравьи организовывали групповую фуражировку и начинался массовый выход фуражиров из гнезд. Работы на поверхности гнезда возобновляются в сентябре, и в ноябре 90% семей ежедневно занимались строительными работами, что явно связано с подготовкой к зимовке. В декабре — феврале все гнезда были закрыты и муравьи на поверхности не появлялись.
За 1970 г. муравьи одного гнезда M. aralocaspius на пробной площадке №1 в среднем собрали 142 г семян, причем 134 г (94%) — в течение апреля и мая. Перед уходом муравьев на зимовку осенью того же года (в октябре и ноябре) было раскопано три гнезда этого вида неподалеку от пробной площадки. Запасы семян в этих гнездах были 0; 12,4 и 13,6 г (среднее 8,7 г). Целесообразность сохранения некоторого количества семян на следующий год несомненна, так как в марте муравьям приходится активно работать, хотя созревших семян в это время еще нет. Впрочем в благоприятные годы запасы могут превосходить потребности семьи. На это указывает тот факт, что при раскопке старых гнезд мы находили замурованные камеры, забитые старыми испорченными у проросшими плесенью семенами.
Численность населения в 5 раскопанных гнездах этого вида была 351, 3343, 5760, 6800 и 11987 особей. Таким образом, за среднюю численность взрослой семьи можно принять 7 тыс. особей, биомасса которых составляет 15 г. Если мы примем NP=B (это, видимо, вполне реальная цифра: у Pogonomyrmex occidentalis в полупустыне Колорадо, США, NP/P=1,01), то экологическая эффективность роста (NP/J) у M. aralocaspius, исходя из наших данных, должна составлять величину порядка 0,1. Это вполне укладывается в пределы, указанные для растительноядных видов (0,08—0,27), но втрое выше, чем у североамериканского муравья-жнеца Pogonomyrmex badius (0,03). На основании материала, изложенного выше, мы построили схему динамики пищевых запасов в гнезде M. aralocaspius, которая изображена на рис:46. Естественно, эта схема не претендует на универсальность, но в общих чертах она, по нашему мнению, отражает общую картину, характерную для облигатного карпофага в условиях одного (весеннего) максимума плодоношения растений.
Еще более контрастную картину представляет сезонная динамика заготовки пищи у M. intermedius на той же пробной площадке. Семьи этого вида пробуждаются от зимней спячки в середине марта. В конце марта они уже начинают собирать старые семена, главным образом днем и в непосредственной близости от гнезд. В апреле большинство семей переходит на ночную активность, и интенсивность фуражировки в двух семьях, находившихся под наблюдением, составляла в среднем около, 20 тыс. экскурсий на гнездо в сутки. В мае среднесуточное количество фуражировочных экскурсий одной семьи увеличилось до 60 тыс. С июня по август продолжался период летнего покоя и муравьи практически не покидали гнезд. В сентябре и октябре они снова вышли из гнезда, но активность в это время была очень низкой — 1,9 тыс. экскурсий на гнездо в сутки. В ноябре начался период зимнего покоя. За 1970 г. население одного гнезда собирало 985 г семян, в том числе 972 г (99%) в апреле и мае. При раскопке крупного гнезда этого вида в Келятинском районе Туркмении в камерах было собрано 1450 г семян.
Иная картина сезонной динамики активности наблюдалась у M. denticulatus (рис:47). Общая продолжительность периода внегнездовой деятельности у этого вида, как и у M. aralocaspius, составляет девять месяцев. Однако в этот период наблюдается гораздо меньшее количество покоящихся муравейников — не более 20—30%. В марте, апреле и мае постоянно активны все семьи, причем, как и у M. aralocaspius, в марте практически все семьи семян не собирают и занимаются ремонтом гнезд. В августе и сентябре ежедневно занимаются фуражировкой 45—65% семей, в октябре их количество возрастает до 80%, а в ноябре падает до 20%. В ноябре снова возрастает (до 70%) количество семей, которые работают только на поверхности гнезд.
Поскольку соотношение активных и покоящихся семей у этого вида изменяется не столь резко, как у M. aralocaspius, динамика средней интенсивности фуражировки и интенсивности фуражировки активных семей сходна. Для M. denticulatus, в отличие от двух других видов, характерны два максимума активности — весенний (апрель—май) и осенний (сентябрь). Различие это, конечно, объясняется различиями в продуктивности растений на пробных площадках: как уже отмечалось выше, на солончаке, где обитает M. denticulatus, наблюдалось осеннее плодоношение растений, отсутствовавшее в пустыне. Минимальная активность у M. denticulatus отмечена в августе. В ноябре и марте муравьи практически не собирали семян. В среднем за год одна семья этого вида на пробной площадке №2 собирала 536 г семян, причем в апреле—мае — 279,5 г (52%) и в сентябре— 186 г (35%).
Параллельно с этой работой был проведен опыт, целью которого было выяснить, сколько семян может запасти семья M. denticulatus при неограниченном количестве корма на территории. Для этого рядом с двумя гнездами этого вида (МД-3, МД-4) на кормовые площадки выкладывалось просо. Размер приманки был таким, что муравьи не могли за сутки перенести корм в гнездо. Приманка постоянно подновлялась и взвешивалась до и после выкладки. Муравьи гнезда МД-3 прекратили работу на пятый день, собрав 468,8 г, а гнезда МД-4 — на восьмой день, собрав 609,8 г семян. После этого муравьи вообще перестали заниматься фуражировкой, несмотря на обилие пищи на кормовом участке. Динамика потребления пищи показана на графике рис:48.
В июне 1971 г. К. Сапарлиев проводил наблюдения за питанием M. denticulatus на полях пшеницы и ячменя в окрестностях Ашхабада до и после уборки. До уборки муравьи собирали главным образом семена сорняков в небольших количествах, но после уборки переключились на сбор потерянного зерна. Наблюдения велись до снижения активности ниже того уровня, который отмечался до уборки. Картина очень напоминала ту, которую мы получили в эксперименте. За девять дней после уборки ячменя (12—20 июня) муравьи собрали по 516,9 г зерна на гнездо (среднее из 10 гнезд). На пшеничных полях за шесть дней (с 22 по 27 июня) после уборки сбор семян составил 721,2 г на гнездо (среднее из десяти гнезд).
Таким образом, двумя независимыми методами были получены очень близкие цифры — 500—700 г/гнездо. Это в 3,5—5 раз больше, чем у M. aralocaspius. Численность и биомасса у M. denticulatus не намного выше, чем у M. aralocaspius, — 8194 экз. и 19,2 г (среднее из трех гнезд, раскопанных в Акыбае). Осенняя раскопка трех гнезд в Акыбае в ноябре 1970 г. показала, что на зиму в гнезде этого вида остается примерно такое же количество семян (0,6; 8,5, 19,4 г, среднее 9,5 г). Правда, у M. denticulatus обнаружено несколько больше крылатых (24,3% биомассы рабочих у M. denticulatus и 14,8% — у M. aralocaspius), но для того, чтобы выкормить этот излишек (2,5 г), при NP/J = 0,1 требуется всего 25 г семян. Как нам кажется, единственное возможное объяснение заключается в том, что M. denticulatus является менее специализированным карпофагом, чем M. aralocaspius, и это заставляет его делать большие запасы при том же размере семьи. Если принять, что у M. denticulatus NP/J=0,03, как у североамериканского Pogonomyrmex badius, то для поддержания постоянной численности и нормального продуцирования крылатых при NP/B=1 семье с биомассой 19 г потребуется 633 г семян, что примерно и наблюдается в естественных условиях. Видимо, этим и объясняется приуроченность M. denticulatus к стациям, богатым пищей (оазисы, солончаки, тугаи), так как строение гнезд и устойчивость к высокой температуре у M. denticulatus и M. aralocaspius существенно не отличаются.
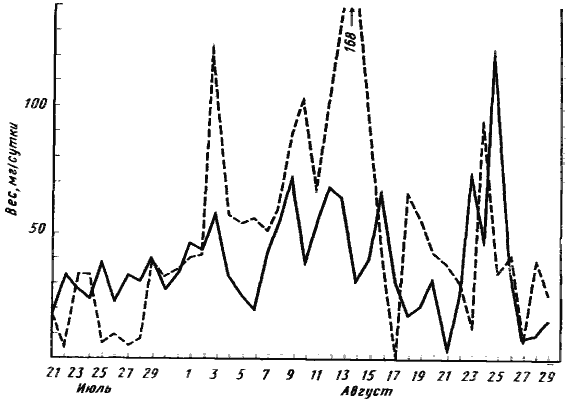
Рис. 43. Динамика потребления белковой пищи (куколки Lasius и Formica) двумя лабораторными семьями
Семьи, состоявшие из 150 рабочих и двух самок одновременно (30 июня 1966 г.), были помещены в одинаковые гнезда. Ежедневно в кормушки выкладывалось одинаковое (избыточное) количество белковой пищи и по остатку определялось количество корма, унесенного за сутки в гнездо. Несовпадение минимумов и максимумов потребления в разных семьях указывает на эндогенные причины колебаний в потреблении
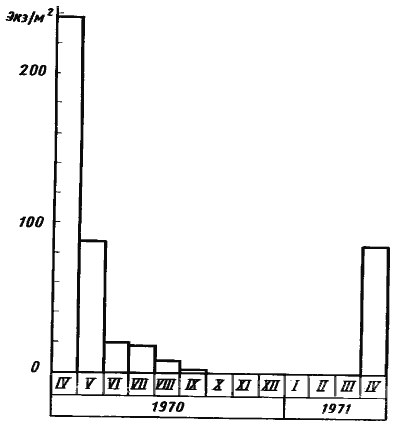
Рис. 44. Сезонная динамика плодоношения эфемеров на песчаных площадках среди такыра в Акыбае в 1970—1971 гг.
Столбики диаграммы — среднее количество растений с созревшими семенами/м2 при одномоментных учетах
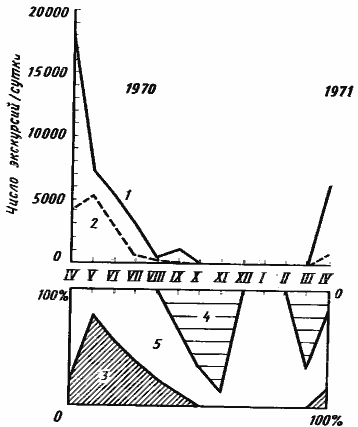
Рис. 45. Сезонная динамика фуражировочной активности Messor aralocaspius в Акыбае в 1970—1971 гг
(по [Длусский, Сапарлыев, 1975], с изменениями)
1 — среднее число фуражировочных экскурсий на активную семью в сутки; 2 — среднее число фуражировочных экскурсий на одну семью (включая покоящиеся) в сутки; 3 — доля (в %) фуражирующих семей; 4 — доля семей, рабочие которых активны на поверхности, но не занимаются фуражировкой; 5 — доля покоящихся семей
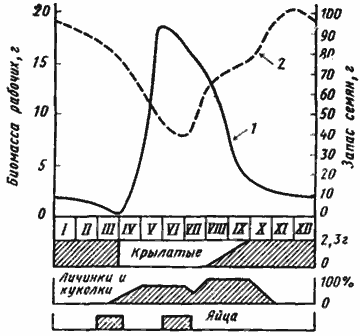
Рис. 46. Обобщенная схема годового жизненного цикла семьи Messor aralocaspius
1 — запас семян в гнезде, г; 2 — биомасса рабочих, сухой вес, г Схема составлена на основании данных раскопок гнезд, учетов приноса семян в гнезда и выноса трупов из гнезд M. aralocaspius в саксаульниках Акыбая в 1970—1971 гг. Средняя численность рабочих в семье принята за 7000 особей, средний сухой вес рабочего — 2,17 мг, B=NP=15,19 г. Гибель в гнезде определена подсчетом выноса трупов в марте — июне 1970—1971 гг. и принята постоянной и равной 16 особей/сут.; гибель на кормовом участке (в основном поедание жужелицей Anthia mannerheimi) принята пропорциональной интенсивности фуражировки. Численность и биомасса расплода оценивалась только ориентировочно, так что кривая отражает только общий характер динамики; за 100% принята максимальная биомасса в августе и сентябре
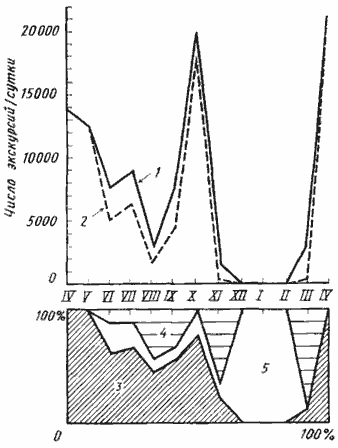
Рис. 47. Сезонная динамика активности Messor denticulatus в Акыбае в 1970—1971 гг.
(по [Длусский, Сапарлыев, 1975], с изменениями)
Обозначения, как на рис.45
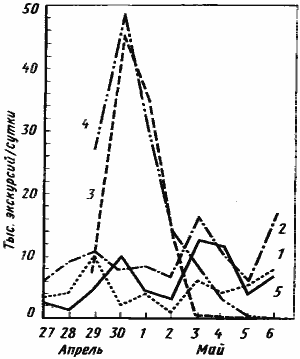
Рис. 48. Влияние избытка пищи на динамику активности Messor denticulatus
(Акыбай, 1970 г.)
1 —5 — номера гнезд (соответственно МД-7001—МД-7005). Рядом с гнездами №3 и 4, 29 апреля были выставлены кормушки с пшеном. Гнезда №1, 2 и 5 служили контролем
(гр. topos - место) - естественное жизненное пространство определенного биоценоза, совместно с которым он образует экосистему.
Всего 134 вида.
Всего 19 видов.
Всего 62 вида.
Одно из крупных подсемейств, насчитывает около 1,5 тыс. видов, относящихся к 44 родам. Широко распространены на всем земном шаре, довольно обычны в тропиках и занимают доминирующее положение в умеренных широтах.
Myrmicinae — самое большое подсемейство муравьев, включающее более 2 тыс. видов, относящихся более чем к 150 родам.
совокупность сложных врожденных реакций (актов поведения) организма, возникающих в ответ на внешние или внутренние раздражения; сложный безусловный рефлекс (пищевой, оборонительный, половой и др.). Инстинкты человека контролируются его сознанием.
ксеробионт
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
участок, используемый муравьями для добычи пищи.
 Myrmica rubra (Linnaeus, 1758) — рыжая мирмика
Myrmica rubra (Linnaeus, 1758) — рыжая мирмикаSubspecies: Myrmica scabrinodis ussuriensis, Kuznetsov-Ugamsky
 Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant
Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden antВсего 965 видов.
необратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.
Старые названия: T. chefteki, T. turcomanica, T. turcmenicum, T. taurocaucasum, T. taurocaucasicum
отряд общественных насекомых. Общины, разделенные на касты, состоят из крылатых и бескрылых особей. Строят подземные и наземные (до 15 м высотой) гнезда (термитники). Ок. 2600 видов, главным образом в тропиках; в России 2 вида: один в районе Сочи, второй, вероятно, завезенный из Японии или Китая, во Владивостоке. Многоядны. Разрушают древесину, портят бумагу, кожу, сельскохозяйственные продукты и др. Термиты, обитающие в почве, полезны как почвообразователи.
животные, не имеющие позвоночника: простейшие, губки, кишечнополостные, черви, моллюски, членистоногие, иглокожие и др.; примерно 3/4 видов всех животных.
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
сладкая густая жидкость, выделяемая тлями, червецами и другими насекомыми, питающимися соком растений. Медоносная пчела собирает падь с листьев и перерабатывает в падевый мед.
организм, питающийся семенами растений.
распространение семян, имеющих съедобные придатки (у фиалки, чистотела и др., главным образом лесных растений), муравьями.
итал. obligato, "обязательный", необходимый, существенный
 Tetramorium caespitum (Linnaeus, 1758) — дерновый муравей, pavement ant
Tetramorium caespitum (Linnaeus, 1758) — дерновый муравей, pavement antСтарое название: T. indocile
Старое название: Pheidole arenarum Ruzsky
 Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушитель
Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушительСтарое название: Monomorium gracillimum
галобионт
Subspecies: Monomorium indicum kusnezowi Santschi, 1928
Старое название: Leptothorax semenovi
Всего 415 видов
(от греч. oikos дом, жилище, местопребывание и ...логия), наука об отношениях живых организмов и образуемых ими сообществ между собой и с окружающей средой. Термин «экология» предложен в 1866 Э. Геккелем. Объектами экологии могут быть популяции организмов, виды, сообщества, экосистемы и биосфера в целом.
общая масса особей одного вида, группы видов или сообщества в целом на единицу поверхности или объема местообитания. Биомасса растений называется фитомассой, биомасса животных зоомассой.
организм, питающийся трупами других животных, гниющими остатками, навозом и т.д.
организм, питающийся растениями
(зоонекрофаг), организм, питающийся мертвыми животными, гетеротроф
организм, питающийся исключительно другими животными
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
Всего 106 видов.
Старое название: Messor rufitarsis
ксеробионт
гигрофил
Всего 60 видов.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
Всего 100 видов.






