






Как показывают данные предыдущего раздела, существенные отличия в термофильности и термостойкости среди пустынных муравьев отмечены лишь у некоторых дневных зоонекрофагов (Acantholepis, Cataglyphis). У карпофагов и ночных зоофагов эти параметры довольно сходны с муравьями-герпетобионтами гумидных областей.
Одним из механизмов повышения термофильности и теплоустойчивости является изменение химического состава белков и связанное с ним изменение их теплостойкости. Такие механизмы были показаны для ряда видов, однако у муравьев до сих пор не изучались. Широко распространено мнение, что различия в окраске покровов у разных форм одного вида насекомых могут быть связаны с различиями в теплоустойчивости. При этом указывается обычно, что в горах и на севере обитает темноокрашенная форма, а на равнине — светлоокрашенная. В частности, такая закономерность была показана для различных видов Formica (Длусский, 1967а, Bernstein, 1976). По крайней мере в одном случае (Тимофеев-Ресовский, Свирежев, 1965) доказана корреляция между окраской и выживанием при различной температуре.
В тех. случаях, когда окраска определяется небольшим числом генов, повышение приспособленности популяции довольно быстро происходит за счет изменения частот этих генов. По-видимому, именно такая картина наблюдается у среднеазиатских Acantholepis. С начала века на территории Средней Азии было описано более десятка подвидов и варьететов A. frauenfeldi, отличающихся окраской и формой чешуйки (Рузский, 1905; Karawajew, 1909; Santschi, 1917; Kuznetzov-Ugamsky, 1929). Впоследствии было обнаружено, что все эти формы довольно сильно отличаются от типичного A. frauenfeldi, распространенного на западе Палеарктики. Некоторые из описанных ранее форм получили видовой статус: так, формы с красной окраской груди были объединены под общим названием A. melanogaster Em., целиком черные — A. karavaievi Sant. и A. spinnisquama K.-Ug. (отличаются формой чешуйки), а формы с промежуточной окраской из Копетдага — под названием A. semenovi Ruzs. (Pisarski, 1967а). Более детальное изучение изменчивости среднеазиатских Acantholepis показало, что в одной популяции и даже одном гнезде могут быть обнаружены разные описанные формы в любых сочетаниях (рис:25/3-5/). При этом в пределах одной популяции соотношение фенотипов в разных гнездах случайно (рис:25/4,5/). Поскольку первой была описана форма с промежуточной окраской (A. frauenfeldi var. semenovi Ruzsky, 1905), приоритет имеет название A. semenovi (syn. nov.), которое и используется в настоящей работе.
В то же время соотношение фенотипов изменяется в пределах ареала вида. Как видно из карты, приведенной на рис:25, в горах обитает исключительно черная форма, а в Каракумах — исключительно красная. В низкогорных и предгорных районах, а также на севере ареала вида (Кызылкумы) встречаются особи с черной, красной и промежуточными окрасками груди, причем все они могут быть обнаружены в одном гнезде. Судя по характеру изменчивости, этот признак скорее всего определяется олигогенной системой. Различие в частотах этих генов в разных популяциях A. semenovi скорее всего связано с тем, что в районах с жарким климатом преимущества получают семьи, в которых самки (вид полигинен) продуцируют светлоокрашенных рабочих, а в более холодных — наоборот.
Интересно, что хотя в Мубареке и в Карши (Каршинская «степь») климат довольно сходен, концентрация генов, обеспечивающих черную окраску, гораздо выше в Карши: в Мубареке даже экземпляры с промежуточной окраской почти не встречаются. Скорее всего это объясняется расстоянием до горных областей, откуда идет постоянный приток генов.
Обратная закономерность наблюдается у Messor aralocaspius. В Бадхызе, в Каракумах, в Каршинской «степи» и на юге Кызылкумов (Шафрикан) все рабочие особи у этого вида имеют черную окраску. В горах в Западном Тянь-Шане и на севере ареала (Кульджуктау, Тартугай) довольно часто встречаются семьи, у которых рабочие имеют красноватую окраску груди (ssp infumatus). Такая же картина наблюдается у Adalia bipunctata в горах Италии (Scali, Creed, 1975).
Многие авторы отмечают, что среди дневных пустынных муравьев встречаются виды со светлой окраской; иногда эта окраска обусловлена густым прилежащим опушением из серебристых волосков (самки Acantholepis, рабочие Cataglyphis bombycina). Несомненно, что такая окраска может быть покровительственной: в песчаной пустыне рабочих Cataglyphis pallida часто можно заметить только по быстро перемещающейся тени животного. Покровительственное значение имеет желтая окраска и для видов с ночной активностью. В пустынях почти никогда не бывает полной темноты и черные муравьи на открытых песчаных или такырных участках довольно хорошо заметны, особенно в лунные ночи. В то же время желтые рабочие совершенно незаметны на фоне почвы даже в сумерках. Однако неверно утверждение, что светлоокрашенные виды характерны именно для пустынной зоны. Скорее наблюдается обратная закономерность. Так, в Каракумах и Кызылкумах 23% видовимеют рабочих с бледно-желтой окраской тела, в лесах умеренной зоны (Московская область) — 30%, а в тропических лесах Тонга — 32%.
Н. Н. Кузнецов (Kusnezov, 1956) высказал предположение, что светлая окраска и густое серебристое опушение служат также для отражения солнечных лучей и защищают их обладателей от перегрева. Для проверки этого предположения мы провели опыт по следующей методике: 2-й тергит брюшка рабочего, очищенный от мягких тканей, помещался между двумя металлическими пластинами, в которых было просверлено отверстие диаметром 1 мм. Второе отверстие такого же диаметра без образца служило контролем. Пластинка помещалась в камеру спектрофотометра СФ-9 и в интервале длины волны 300—1150 нм снимался спектр пропускания кутикулы 2-го брюшного тергита. Предварительно диаметры отверстий были отъюстированы, так что без образцов они пропускали одинаковое количество света.
В качестве объектов были взяты Camponotus turcestanicus, Cataglyphis setipes и C. pallida из Юго-Западных Кызылкумов. Первый вид ведет ночной образ жизни, рабочие второго и третьего активны днем. Окраска первых двух видов довольно сходна — красная грудь и буровато-черное брюшко. C. pallida целиком бледно-желтого цвета. У всех трех видов брюшко покрыто тонкой поверхностной скульптурой и редкими прилежащими волосками.
Результаты измерений оказались несколько неожиданными. Как видно из графиков на рис:26, несмотря на сходство окраски брюшка C. setipes и C. turcestanicus, спектры пропускания кутикулы у этих видов оказались весьма различными. В то же время оптические свойства кутикулы C. setipes и C. pallida, сильно отличающихся по окраске, во многом сходны. Для ультрафиолетовых лучей кутикула всех трех видов почти непрозрачна. В области 300—600 нм (ультрафиолетовый — желтый) кутикула у C. pallida более проницаема, чем у C. setipes, но в длинноволновой части спектра наблюдается обратная картина. Особенно заметны различия в инфракрасной области (780—1150 нм): кутикула у C. pallida пропускает всего 6,8—9,2% излучения, а у C. setipes — от 10,2% при длине волны 780 нм до 21% при длине волны 1150 нм. Кутикула C. turcestanicus оказалась более прозрачной во всех областях изученного спектра. При длине волны 700 нм (красный цвет) и выше пропускается около 50% света. По всей видимости, это свидетельствует о том, что сама кутикула прозрачна в этой части спектра, а 50% энергии рассеивается за счет структурированности ее поверхности.
На основании полученных данных можно, видимо, говорить, что у пустынных дневных герпетобионтов-зоонекрофагов, во всяком случае у некоторых видов, кутикула значительно менее прозрачна в длинноволновой части спектра, чем у ночных зоофагов, имеющих для нашего глаза сходную окраску. Эти различия скорее всего обусловливаются составом пигментов, так как структура кутикулы изученных видов в принципе довольно сходна, а толщина ее у C. pallida даже меньше, чем у C. setipes.
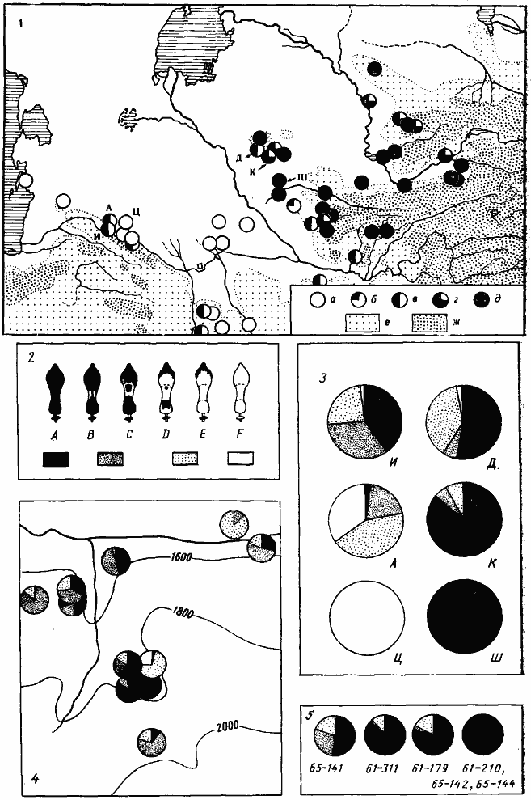
Рис. 25. Изменчивость окраски у Acantholepis semenovi
1 — географическое распространение A. semenovi и его изменчивость в пределах ареала:
а — только особи с красной грудью и чешуйкой (тип F);
б — главным образом красная форма, но встречаются более темные особи (типы D—F);
в — встречаются все варианты окраски;
г — главным образом черные особи, с некоторой примесью светлоокрашенных (типы А—D);
д — только черная форма (из некоторых районов имеются лишь небольшие выборки);
е — участки, лежащие на высоте 500—2000 м;
ж — участки на высоте более 2000 м;
2 — варианты окраски груди и обозначения этих вариантов на круговых диаграммах;
3 — соотношение разных фенотипов в 6 популяциях (обозначения те же, что на карте):
И — Ипай-Кала, Копетдаг, высота 1500—2000 м;
А — Арчман, предгорья Копетдага;
Ц — Каррыкуль, Центральные Каракумы;
Д — Джингельды, Юго-Западные Кызылкумы, в 30 км восточнее отрогов Кульджуктау;
К — Аяк-Гужумды, предгорья Кульджуктау;
Ш — Шафиркан (небольшая изолированная мономорфная популяция в саксауловом лесу);
4 — соотношения
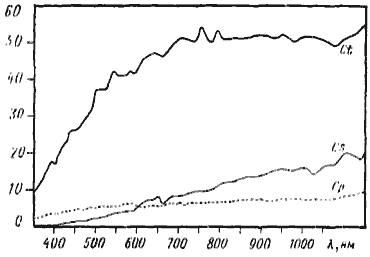
Рис. 26. Спектры пропускания кутикулы 2-го брюшного тергита у Camponotus turcestanus (Ct), Cataglyphis setipes (Cs) и C. pallida (Cp)
(зоонекрофаг), организм, питающийся мертвыми животными, гетеротроф
Всего 65 видов.
организм, питающийся семенами растений.
организм, питающийся исключительно другими животными
организм, обитающий среди растительных или иных органических остатков, на поверхности почвы.
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
совокупность особей одного вида, длительно занимающая определенное пространство и воспроизводящая себя в течение большого числа поколений. В современной биологии популяция рассматривается как элементарная единица процесса эволюции, способная реагировать на изменения среды перестройкой своего генофонда.
Старое название: Acantholepis frauenfeldi
Старые названия: Acantholepis semenovi, Acantholepis transcaspiensis
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
наличие в семье нескольких яйцекладущих самок. Смотри: моногиния, олигогиния.
ксеробионт
Старое название: Cataglyphis bombycina
сегмент брюшка.
ксеробионт






