






Для зоонекрофагов, обладающих морфофизиологическими адаптациями к перенесению высокой температуры и низкой влажности (Acantholepis, Cataglyphis), характерен дневной тип активности. В качестве примера можно привести суточный цикл фуражировочной активности семьи Cataglyphis setipes №71—30 9 мая 1971 г. (рис:38). Первые рабочие появились из этого гнезда в 8 ч утра, т.е. через 2 ч после восхода солнца, когда температура воздуха поднялась до 18,5°, а температура почвы — до 19,5°. Муравьи расходились во всех направлениях от гнезда. В 8 ч 45 мин первые фуражиры начали возвращаться, причем все они несли добычу, главным образом трупы насекомых. Их возвращение стимулировало новый выход из гнезда. Часть вышедших муравьев не уходила далеко от гнезда и вскоре возвращалась в него без всякой добычи. В 10 ч фуражиры гнезда Messor aralocaspius, находившегося поблизости, покинули территорию. Это вызвало новую волну выхода: фуражиры C. setipes вышли на территорию, где перед этим работали жнецы, и начали собирать трупы муравьев-жнецов, погибших за ночь. По мере возвращения фуражиров с трупами Messor активность начала постепенно спадать и достигла минимума к 12 ч. В 13 ч гнездо, которое до этого находилось в тени саксаула, оказалось на солнце. Температура его поверхности за короткое время поднялась с 30 до 56°. Это вызвало новую волну выхода. Но поведение муравьев на кормовом участке изменилось. Часть муравьев, выйдя из гнезда, направились в кроны саксаулов. Большинство же выбегали из гнезда, с большой скоростью обегали участок территории и быстро возвращались обратно независимо от того, удалось им обнаружить добычу или нет. Естественно, резко возрос процент особей, не несущих добычи. После 16 ч температура на почве начала понижаться и одновременно начала снижаться активность муравьев. К 18 ч прекратился выход из гнезда, несмотря на то, что температура воздуха была 29°, а температура почвы вблизи гнезда — 44° и до захода солнца оставалось еще около 2 ч. К 19 ч в гнездо вернулись последние рабочие.
Из приведенных данных видно, что высокая температура (более 50— 60° на почве) не служит препятствием для фуражировки C. setipes. В интервале температур на поверхности почвы 20—70° внегнездовая активность определяется освещенностью, наличием пищи и динамикой температуры (повышение, понижение). Еще более теплолюбивыми видами являются Cataglyphis pallida и Acantholepis semenovi (красная форма), которые начинают фуражировку позже и заканчивают раньше, чем C. setipes.
Совсем иная картина фуражировки наблюдается у большинства видов муравьев-жнецов, не обладающих морфофизиологическими приспособлениями к высокой температуре и низкой влажности, но в то же время не способных ориентироваться в темноте (Длусский, Сапарлыев, 1975), и у Monomorium barbatulum. В качестве примера можно рассмотреть суточный ритм внегнездовой активности у Messor denticulatus (рис:39/2,3/). В апреле первые муравьи появляются из гнезд в 7—8 ч, а в мае — августе — в 4—5 ч, т.е. с началом рассвета. Сразу же из гнезда появляется большое количество рабочих, которые через 10—20 мин после выхода формируют тропу к участку сбора семян. Вскоре после начала массового выхода из гнезда наблюдается, как и у C. setipes, некоторый спад интенсивности выхода, а затем она снова поднимается и стабилизируется. Интенсивность возвращения в это время поднимается медленнее, и оба потока фуражиров становятся одинаковыми лишь через 1—2 ч после начала внегнездовой активности. Когда температура на поверхности почвы достигает 32—35°, интенсивность выхода из гнезда M. denticulatus начинает падать. При дальнейшем повышении температуры муравьи начинают интенсивно покидать кормовой участок и возвращаться в гнездо. В это время резко возрастает процент муравьев, возвращающихся в гнездо без пищи. При температуре поверхности почвы 37—45° внегнездовая деятельность муравьев полностью прекращается. Во второй половине дня, когда температура падает до 32—37°, муравьи начинают снова выходить из гнезд. Как правило, сразу же образуется дорога. Когда начинает темнеть, интенсивность выхода из гнезда падает и муравьи начинают возвращаться. С наступлением темноты всякая фуражировка прекращается, однако обмен муравьями между разными гнездами по уже стабилизировавшейся тропе может продолжаться и ночью.
Важно отметить, что у всех видов с утренне-вечерним ритмом активности дневной перерыв фуражировки является вынужденным и обусловливается только высокой температурой. В тех редких случаях, когда температура поверхности почвы в середине дня не превышает 32—35°, такого перерыва не бывает и максимум активности наблюдается днем. Температура на поверхности почвы обусловливает и длительность дневного перерыва (рис:39/1/).
Утренне-вечерний тип активности изучен также у Messor aegyptiacus из Сахары (Sheata, Kaschef, 1972) и муравьев-жнецов пустынь Северной Америки — Veromessor pergandei (Went, Wheeler, Wheeler, 1971; Clark, Comanor, 1973), Pogonomyrmex spp. (Whitford, Ettershank, 1975; Hansen, 1978), разводящего гриба пустынного Trachymyrmex smithi neomexicanus (Schumacher, Whitford, 1974). Имеются указания на наличие такой ритмики у многих европейских и африканских Messor и Monomorium chobauti (Sudd, 1967; Delye, 1968).
Поскольку перерыв фуражировочной активности в середине дня является у всех муравьев вынужденным, утренне-вечерний тип активности в принципе не отличается от дневного типа, характерного для зоонекрофагов. Все различия в конце концов сводятся к различиям в устойчивости к высокой температуре и низкой влажности. Для дневных зоонекрофагов пустынь Средней Азии верхняя температурная граница активности лежит выше 60°, так что спад активности в середине дня удается наблюдать редко. У Conomyrma insana в Северной Америке снижение интенсивности фуражировки отмечается при температуре выше 57° (Van Pelt, 1966). Сахарские Cataglyphis bombycina и C. albicans активны при температуре 58°, но уже при 50° (на поверхности почвы) у них наблюдается некоторое снижение активности. У Cataglyphis bicolor и Acantholepis frauenfeldi Сахары этот порог несколько ниже — 46—47° (Delye, 1968). В подгорной степи Зеравшанского хребта (Варганза) мы наблюдали перерыв фуражировки у дневных зоонекрофагов Proformica epinotalis и Acantholepis semenovi (черная форма) при повышении температуры почвы выше 50°. Примерно такие же пороговые температуры указываются для североамериканских пустынных Pogonomyrmex: максимальное число рабочих, возвращающихся в гнездо, отмечено при 45°; при температуре 50° почти все фуражиры покидают территорию, хотя отдельные особи могут возвращаться (не погибают) при 65° (Whitford, Ettershank, 1975). У Veromessor pergandei рабочие покидают территорию при повышении температуры поверхности почвы более 39° (Clark, Comanor, 1973). В Сахаре у Messor caviceps, M. arenarius и Monomorium chobauti снижение активности происходит при температуре 41—42°, а у Messor aegyptiacus — 38—39° (Delye, 1968). У дневных герпетобионтов умеренной зоны, например у Formica pratensis, дневной спад активности начинается, когда поверхность почвы нагревается до 30—32° (Длусский, 1967а).
Следует, однако, отметить, что к сравнению цифр, полученных разными авторами в разных условиях, нужно подходить с большой осторожностью. Дело в том, что температурные пределы фуражировочной активности не являются абсолютным свойством вида и могут в широких пределах (приблизительно 3—5°) варьировать у особей одной и той же семьи в зависимости от привыкания к высокой (или низкой) температуре. Так, например, по нашим наблюдениям, в Акыбае у M. aralocaspius и M. denticulatus в марте выход муравьев из гнезд после дневного перерыва начинался при температуре 32—33°, а летом рабочие из тех же семей начинали выходить для фуражировки при 37—38°. Сходные данные приводит Грэй (Gray, 1971с) для двух австралийских Myrmecia. У M. desertorum рабочие активны летом при температуре поверхности почвы 19—48° и зимой — 10—42°, а у M. dispar — соответственно 19—44° и 10—35°. Таким образом, имеющиеся данные позволяют лишь заключить, что у специализированных пустынных зоонекрофагов дневной перерыв активности начинается при повышении температуры почвы более 55°, у неспециализированных пустынных зоонекрофагов, зоонекрофагов семиаридных областей и специализированных дневных фитофагов пустынь — 45—55°, у неспециализированных к перенесению высокой температуры дневных карпофагов — 35—45° и у дневных зоонекрофагов умеренной зоны — ниже 35°.
В отличие от видов, описанных выше, некоторые пустынные виды муравьев способны ориентироваться и вести фуражировку ночью. Это дает им в условиях пустыни огромные преимущества. Так, по нашим данным (Длусский, Сапарлыев, 1975), в Южной Туркмении рабочие Messor aralocaspius в апреле — мае (период созревания семян эфемеров) в среднем могут работать ежедневно 6 ч 30 мин (4 ч 15 мин утром и 2 ч 15 мин вечером). В тех же условиях M. intermedius, у которого отсутствуют морфофизиологические приспособления к перенесению высокой температуры (верхняя граница активности 30—35°, как у муравьев гумидных районов умеренной зоны), только за счет способности вести фуражировку в темноте работают ежедневно в среднем 12 ч 30 мин, т.е. почти вдвое больше. А это при том же количестве фуражиров позволяет за то же время сделать в 1,5—2 раза большие запасы семян.
В пустынях Средней Азии мы наблюдали ночную фуражировку у Tetramorium striativentre, T. inerme, T. nitidissimum, Pheidole arenarum, Monomorium kusnezovi, Leptothorax semenovi, Messor intermedius, Messor группы excursionis (M. variabilis, M. excursionis, M. lamellicornis) и всех пустынных видов Camponotus. В литературе имеются указания на ночную активность Messor barbarus (?) и Camponotus thoracicus в пустынях Северной Африки (Sudd, 1967; Delye, 1968), Pogonomyrmex rugosus, Novomessor cockerelli (Whitford, Ettershank, 1975), N. manni (Kannowski, 1954) и некоторых видов Myrmecocystus (McCook, 1881; Wheeler, 1908; Schumacher, Whitford, 1976, 1978) в пустынях Северной Америки. По наблюдениям Н. Н. Дроздова и А. А. Захарова (личн. сообщ.), в пустынях Австралии ночью активны некоторые крупные Rhytidoponera, Bothroponera, Camponotus и некоторые Iridomyrmex.
Детали биологии ночных видов изучены пока еще очень плохо. Это связано с трудностями наблюдения: при освещении нормальное поведение этих муравьев нарушается и они либо стремятся скрыться, либо, наоборот, движутся к фонарю. При изучении видов с крупными рабочими (Messor intermedius, Camponotus xerxes) нам удалось преодолеть эту трудность, освещая территорию слабым светом с большого расстояния (лучше, если фонарь зажжен до наступления темноты) или освещая муравьев короткими вспышками электрического фонаря через значительные интервалы. Наблюдения облегчало и то, что выбирались гнезда на такыровидных площадках, лишенных растительности. Однако за видами, рабочие которых имеют мелкие размеры, при слабом свете наблюдать невозможно, так что сведений о их поведении ночью собрать почти не удалось.
У Tapinoma karavaievi, Monomorium kusnezovi, ![]() M. gracillimum, Pheidole arenarum и всех видов Tetramorium рабочие могут быть активны днем, в пасмурную погоду или в тени саксаулов, солянок и кустиков полыни. Максимум активности этих видов приходится на вечерние и утренние часы, когда температура почвы ниже 30—32° (рис:7). Но с наступлением темноты, в отличие от Messor denticulatus или Monomorium barbatulum, их активность хотя и снижается, но полностью не прекращается, если температура не опускается ниже 10—15°.
M. gracillimum, Pheidole arenarum и всех видов Tetramorium рабочие могут быть активны днем, в пасмурную погоду или в тени саксаулов, солянок и кустиков полыни. Максимум активности этих видов приходится на вечерние и утренние часы, когда температура почвы ниже 30—32° (рис:7). Но с наступлением темноты, в отличие от Messor denticulatus или Monomorium barbatulum, их активность хотя и снижается, но полностью не прекращается, если температура не опускается ниже 10—15°.
Истинно ночная активность наблюдается у пустынных Camponotus, Leptothorax группы semenovi и Messor группы excursionis. Эти муравьи появляются на поверхности почвы только после захода солнца и большинство их рабочих до восхода солнца возвращается в гнезда. Особенно ярко выражена специализация к ночному образу жизни у Camponotus, и в частности у C. xerxes (рис:40). У этого вида рабочие начинают выходить из гнезда вскоре после захода солнца, и за сравнительно короткое время (40—50 мин) гнездо покидают все фуражиры. Из одной секции гнезда с 500—600 рабочих выходит около 130 фуражиров, в том числе около 90 трофобионтов и около 40 охотников. Трофобионты отправляются на ближайшие колонии тлей или червецов для сбора пади, а охотники расходятся по сектору (ширина его зависит от числа секций в гнезде) и уходят на расстояние более 50 м. Приблизительно через час после начала активности в гнездо начинают возвращаться первые трофобионты. В среднем они совершают за ночь по два рейса. Охотники совершают только по одному рейсу, причем почти все они возвращаются с добычей. Фуражировка прекращается вскоре после восхода солнца, хотя мне приходилось видеть отдельных возвращавшихся фуражиров в 8—9 ч утра. Очень сходную картину фуражировки описывает Мак-Кук (МсСоэк, 1881) у североамериканского Myrmecocystus hortusdeorum.
У Messor из группы excursionis и Leptothorax semenovi (рис:12) выход рабочих также начинается после захода солнца, почти в темноте. Однако у этих зерноядных видов он растянут и муравьи продолжают покидать гнездо всю первую половину ночи. Рабочие расходятся из гнезд поодиночке и занимаются сбором семян. С наступлением рассвета фуражировка прекращается. У всех ночных видов активная внегнездовая деятельность происходит при температуре 10—30°, что почти совпадает с цифрами для большинства видов гумидных областей умеренной зоны.
До сих пор остается неясным, как эти муравьи, фуражируя поодиночке, находят обратную дорогу к гнезду. Ни Messor variabilis, ни Camponotus xerxes, уходя из гнезда, не оставляют пахучих следов. Рабочие C. xerxes, разбегающиеся от гнезда, держат брюшко даже слегка приподнятым вверх. Судя по всему, светочувствительность зрения муравьев существенно не отличается от нашей (Мазохин-Поршняков, Тренн, 1972). Можно было бы предположить, что муравьи используют астронавигацию по луне, но, как показали специально проведенные исследования, у ночного Camponotus maculatus отсутствует корреляция между активностью и фазами луны (Kaschef, Sheata, 1963). Как мне кажется, логичнее всего предположить, что муравьи запоминают ольфакторный ландшафт местности (запаховые пятна и поля). Это, кажется, подтверждается и небольшими наблюдениями, которые удалось сделать осенью 1971 г. в Акыбае. На такыре, где было расположено гнездо C. xerxes, было насыпано несколько площадок из эолового песка с вершин барханов, на котором крупные муравьи оставляют хорошо заметные следы. Каждое утро на этих площадках находили не только обычные следы ног C. xerxes, но и короткие (около 10 см) извилистые бороздки, оставленные брюшком (характерный след при мечении субстрата феромоном). Поскольку все дни наблюдения семья не проводила мобилизаций в зону с площадками, можно предположить, что оставленные пахучие следы — это «запаховые маяки». Кстати, оставление «запаховых маяков» очень часто можно наблюдать у многих видов муравьев в лабораторных условиях, когда муравьи осваивают новую арену.
Особое место среди муравьев пустынь Средней Азии занимает Messor intermedius, самый высокоорганизованный вид муравьев среднеазиатских пустынь. В Акыбае в конце марта и начале апреля эти муравьи обычно активны днем. Но с середины апреля, когда в массе созревают эфемеры и одновременно значительно поднимается температура в середине дня, они переходят к ночной активности. Ночную активность мы наблюдали у этого вида в Центральных Каракумах (Каррыкуль) в первой половине апреля 1970 г. В период массового сбора семян (рис:41) выход из гнезда начинается вскоре после захода солнца, когда температура на почве падает до 35°. Сразу же устанавливается дорога до кормового участка. Численность фуражиров у M. intermedius значительно больше, чем у M. aralocaspius или M. denticulatus, и в первую половину ночи, когда активность M. intermedius максимальна, по дороге средней семьи ежеминутно проходит в одном направлении 100—200 особей. После полуночи поток муравьев из гнезда начинает ослабевать, и к рассвету выход полностью прекращается. Возвращение в гнездо продолжается 2—3 ч после восхода солнца.
Однако описанное поведение нельзя считать типичным для этого вида. В более влажных районах (пустоши в окрестностях Ашхабада, Бадхызский заповедник), где выше и плотность гнезд и численность населения в одном гнезде, этот муравей имеет утренне-вечерний тип активности, характерный для большинства палеарктических муравьев-жнецов. В Акыбае мы также наблюдали одну небольшую (очевидно молодую) семью этого вида, которая работала утром и вечером.
Привлекает внимание тот факт, что при ночной фуражировке M. intermedius мы никогда не наблюдали работы фуражиров вне основной дороги и основного поля, на котором собирались семена. Образование дороги является обязательным условием ночной фуражировки. В то же время рабочие упоминавшейся выше небольшой семьи, работавшие на свету, в отдельные дни собирали семена не образуя дороги; фуражиры при этом расходились поодиночке во всех направлениях от гнезда. Так же ведут себя и рабочие из больших семей ранней весной, когда они фуражируют днем.
Все это заставляет предположить, что рабочие M. intermedius не способны ориентироваться в темноте при одиночной фуражировке, как это делают Messor из группы excursionis. Однако при групповой фуражировке поток муравьев на дороге, сформировавшейся еще до наступления темноты, служит хорошим ориентиром для выхода на кормовой участок и возвращения в гнездо. Возможно, какую-то роль играет и акустическая ориентация, так как звук челюстей массы рабочих Messor intermedius, работающих на поле созревших эфемеров, слышен ночью за несколько метров. Выкладывая вечером обильную приманку, можно заставить работать ночью и M. denticulatus. В этом случае ориентация муравьев также происходит по потоку фуражиров.
Похожая корреляция между типом фуражировки и ритмикой суточной активности описана также у североамериканских муравьев-жнецов из рода Pogonomyrmex в пустыне Чиуауа (Whitford, Ettershank, 1975). P. rugosus, в семьях которого насчитывается 1—6 тыс. фуражиров, образует при фуражировке дороги и работает ночью, тогда как у P. californicus (около 1000 фуражиров) и P. desertorum (200—600 фуражиров) фуражиры работают поодиночке и сбор семян происходит только днем. По наблюдениям Ханта (Hunt, 1977), Veromessor pergandei, считавшийся дневным видом, в пустынях летом часто активно фуражирует ночью, но происходит это только в лунные ночи. В безлунную погоду либо фуражировка не происходит, либо дороги плохо организованы.
Как это ни странно, но способность муравьев работать ночью (за исключением случаев типа M. intermedius) не вырабатывается у муравьев как специальная адаптация к жизни в пустыне. Мак-Класки (McClus-key, 1973, 1974) обратил внимание, что характер ритмики активности сходен у близких родов муравьев. Действительно, Formica гумидных областей (исходный род для трибы Formicini) — типичные дневные виды. И все роды трибы, сформировавшиеся и эволюировавшие в семиаридных и аридных областях (Proformica, Alloformica, Cataglyphis), также имеют дневную активность и неспособны ориентироваться ночью. Напротив, для Lasius гумидных областей характерна либо ночная активность (геобионты), либо круглосуточная активность с ночным максимумом. Соответственно некоторые Myrmecocystus североамериканских пустынь, относящиеся к той же трибе Lasiini, несмотря на внешнее сходство с Cataglyphis, фуражируют ночью и занимают ту же экологическую нишу, что и Camponotus палеарктических пустынь.
Для многих Camponotus гумидных и семиаридных областей характерна либо ночная, либо круглосуточная активность с ночным максимумом. Типично ночная активность наблюдается, например, у европейского C. nylanderi (Baroni Urbani, 1965). У обычного в наших северных лесах C. herculeanus рабочие активны круглые сутки, причем движение на дороге прекращается ранним утром, несколько увеличивается с подъемом температуры, а затем снова уменьшается в середине дня. Вечером активность муравьев начинает возрастать и достигает максимума в середине ночи. У C. japonicus в южном Приморье (Уссурийский заповедник) основная масса фуражиров выходит из гнезда с наступлением темноты и возвращается в гнездо утром, хотя отдельные рабочие занимаются фуражировкой днем. Причем такая циклика наблюдается даже в очень влажную погоду (во время наблюдений в конце августа 1968 г. в течение двух недель почти непрерывно шел дождь). Поэтому не удивительно, что для большинства Camponotus пустынь Палеарктики, Южной Африки и Австралии характерна ночная активность.
Среди Aphaenogaster, которые скорее всего являются предками Messor, встречаются как виды с выраженной ночной активностью (A. subterranea), так и дневные виды (A. raphidiiceps). Также и среди Messor встречается группа видов, способных активно ориентироваться ночью (группа M. excursionis). Кстати, возможно этот факт указывает на независимое происхождение группы M. excursionis от особой группы Aphaenogaster.
Подводя итоги этого раздела, можно отметить, что среди пустынных муравьев выделяется две достаточно обособленные группы видов — виды, способные и неспособные ориентироваться в темноте. Представители первой группы получают возможность использовать для фуражировки наиболее благоприятное ночное время. Однако способность к ночной фуражировке возникает у муравьев еще в гумидных областях и при переходе к жизни в пустыне может только усиливаться. Виды, не способные активно ориентироваться в темноте, в условиях пустыни либо вырабатывают морфофизиологические адаптации к высокой температуре и низкой влажности, либо переходят к вынужденной утренне-вечерней активности. И лишь немногие наиболее высокоорганизованные виды этой группы (M. intermedius) вырабатывают в условиях пустыни особую стратегию фуражировки, позволяющую им собирать пищу ночью.
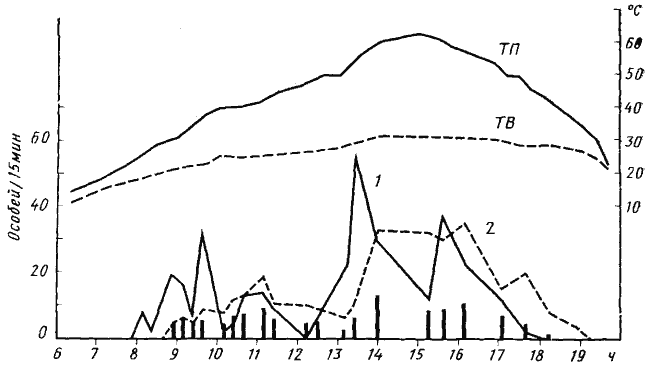
Рис. 38. Суточная динамика фуражировочной активности Cataglyphis setipes
(Акыбай. 10 мая 1971 г., гнездо №71-30)
ТВ — температура воздуха в тени на высоте 1 м; ТП — температура поверхности почвы на солнце (до 13 ч 15 мин вход в гнездо находился в тени саксаула и температура на поверхности почвы вблизи него была приблизительно равна температуре воздуха в тени);
1 — интенсивность выходов из гнезда (выходов/15 мин); 2 — интенсивность возвращения в гнездо; вертикальные столбики — число единиц добычи, принесенных в гнездо за 15 мин
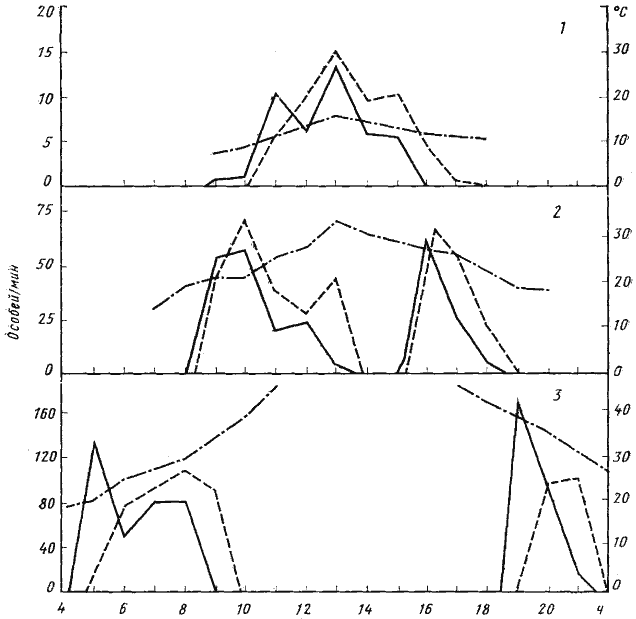
Рис. 39. Суточная динамика фуражировочной активности Messor denticulatus при различной температуре поверхности почвы
(Акыбай) [Длусский, Сапарлыев, 1975]
1 — гнездо МД-705, 24 марта 1971; 2 — гнездо МД-703, 21 апреля 1971 г.; 3 — гнездо МД-703, 2 июля 1971 г.
Сплошная линия — интенсивность выхода из гнезда (особей/мин); пунктир — интенсивность возвращения; штрихпунктир — температура на поверхности почвы
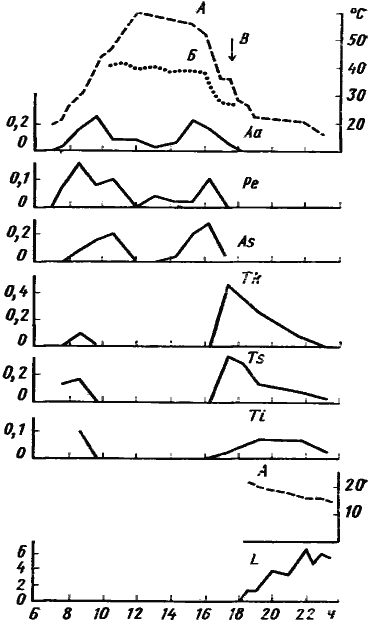
Рис. 7. Динамика суточной активности муравьев-герпетобионтов в подгорной степи
(Варганза)
По вертикали — средняя динамическая плотность (особей/дм2/мин) или (L. semenovi) число особей а квадрате 10x10 см с входом в центре. А — температура на поверхности почвы; Б — температура в траве на высоте 4 см над землей; В — время захода солнца Aa — Alloformica aberrans; Ре — Proformica epinotalis; As — Acantholepis semenovi; Tk — Tapinoma karavaievi; Ts — Tetramorium striativentre; Ti — Tetramorium inerme; L — Leptothorax semenovi
Нижний график — 27 мая, остальные — 22 мая 1978 г.
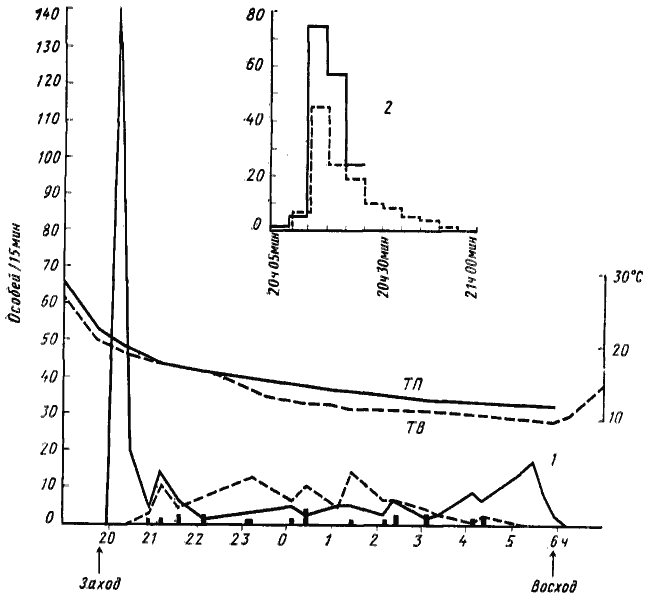
Рис. 40. Суточная динамика фуражировочной активности Camponotus xerxes
(Акыбай, 10—11 мая 1971 г., гнездо № 71-31)
1 — суточная динамика активности 10—11 мая (обозначения, как на рис.38); 2 — развертка динамики первого выхода вечером 11 мая. Сплошная линия — число особей, вышедших из гнезда; пунктир — число особей, поднявшихся на саксаул с колониями тлей и червецов. Замеры по 5 мин, подсчет велся непрерывно. Выход из гнезда кончился в 20 ч 50 мин, а подъем на дерево — в 20 ч 55 мин. В 21 ч 00 мин начали возвращаться первые трофобионты. Всего за это время из гнезда вышла 131 особь, в том числе 97 трофобионтов
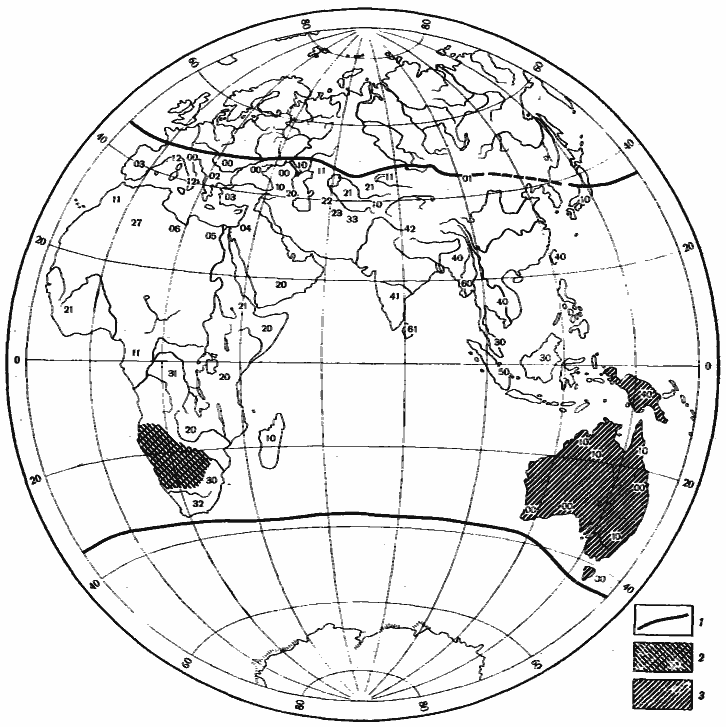
Рис. 12. Некоторые закономерности распространения Camponotus в Восточном полушарии
1 — границы распространения подрода Tenaemyrmex; индекс в пределах ареала подрода: первая цифра — число видов группы C. compressus—sylvaticus (в том числе C. xerxes и C. turcestanus), вторая — группы C. irritans—thraso (в том числе C. turcestanicus и C. fedtschenkoi); 2 — распространение подрода Myrmopsamma; 3 — распространение подрода Myrmophyma (по данным [Emery, 1920] с дополнениями)
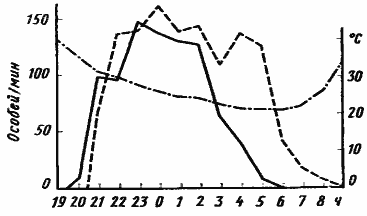
Рис. 41. Суточная динамика фуражировочной активности Messor intermedius
(Акыбай, 17—18 мая 1970 г., гнездо МИ-12) [Длусский, Сапарлыев, 1975]
Обозначения, как на рис.39
(зоонекрофаг), организм, питающийся мертвыми животными, гетеротроф
Всего 65 видов.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
ксеробионт
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
ксеробионт
Всего 106 видов.
участок, используемый муравьями для добычи пищи.
Старые названия: Acantholepis semenovi, Acantholepis transcaspiensis
гигрофил
Старое название: Novomessor pergandei, Veromessor pergandei
Всего 60 видов.
Старое название: Conomyrma insana
Старое название: Cataglyphis bombycina
Старое название: Acantholepis frauenfeldi
организм, обитающий среди растительных или иных органических остатков, на поверхности почвы.
Всего 90 видов.
организм, питающийся растениями
организм, питающийся семенами растений.
Старое название: Pheidole arenarum Ruzsky
галобионт
Subspecies: Monomorium indicum kusnezowi Santschi, 1928
Старое название: Leptothorax semenovi
Всего 965 видов.
Старое название: Messor rufitarsis
Старое название: Novomessor cockerelli
Старое название: Novomessor manni
Всего 30 видов.
Всего 114 видов.
Всего 58 видов.
гигрофил, галобионт
 Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушитель
Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушительСтарое название: Monomorium gracillimum
Всего 415 видов
Всего 19 видов.
взаимовыгодные отношения между муравьями и выделяющими падь насекомыми: слизывая падь, муравьи охраняют этих насекомых.
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
общее название нескольких семейств насекомых подотряда кокцидовых. Св. 1600 видов, преимущественно в тропиках и субтропиках. У тропических лаковых червецов покровы образованы застывающими на воздухе лаковыми выделениями. Многие (напр., мучнистые червецы) вредители растений, особенно субтропических (напр., цитрусовых).
сладкая густая жидкость, выделяемая тлями, червецами и другими насекомыми, питающимися соком растений. Медоносная пчела собирает падь с листьев и перерабатывает в падевый мед.
часть гнезда рыжих лесных муравьев, занимаемая одной колонной.
Старsые названия: M. hortideorum, M. hortusdeorum
(греч. φέρω — «нести» + ορμόνη — «гормон») — летучие вещества, управляющие поведенческими реакциями, процессами развития, а также многими процессами, связанными с социальным поведением и размножением. Феромоны модифицируют поведение, физиологическое и эмоциональное состояние или метаболизм других особей того же вида.
индивидуальное или групповое привлечение пассивных фуражиров активными фуражирами для конкретных полей в конкретном месте.
Всего 24 вида.
Всего 4 вида.
Всего 100 видов.
организм, обитающий на поверхности и в толще пород суши.
(от греч. oikos дом, жилище, местопребывание и ...логия), наука об отношениях живых организмов и образуемых ими сообществ между собой и с окружающей средой. Термин «экология» предложен в 1866 Э. Геккелем. Объектами экологии могут быть популяции организмов, виды, сообщества, экосистемы и биосфера в целом.
ночная активность
Всего 172 вида.
Включает сокращенный Novomessor.
интенсивность посещения муравьями единицы площади в единицу времени (особь/дм²хмин.)






