






При анализе фауны пустынь бросается в глаза большое по сравнению с лесной зоной число фоновых видов. Под фоновыми или обычными я здесь понимаю виды, гнезда которых встречаются по крайней мере на каждые 2 км маршрута для крупных или на каждой из 10 площадок 10х10 м для мелких муравьев. Так, в саксауловых лесах дельты Мургаба (Акыбай) фоновыми являются 16, а в грядово-такырном комплексе Центральных Каракумов 12 видов (Длусский, 19746, 1975а). Это довольно много: достаточно сказать, что в лесах Южного Приморья, где нами обнаружено около 25 видов муравьев, фон составляют пять — шесть видов (Длусский, Купянская, 1972а). При этом следует учитывать, что в лесах муравьи могут использовать гораздо большее число ярусов, а в пустыне все действие развертывается в одной плоскости. Согласно классификации биоморф (жизненных форм) муравьев, предложенной К. В. Арнольди (1937, 1968а), все виды муравьев, обитающих в пустыне, за исключением дендробионтов Crematogaster subdentata и Camponotus semirufus, связанных с саксаулом, Solenopsis deserticola, обитающего в термитниках, и Leptanilla alexandri, живущей, по-видимому, в норах песчанок (Длусский, 19696), относятся к двум биоморфам — герпетобионтов-фитофагов и герпетобионтов-зоофагов. Все они добывают пищу главным образом на поверхности почвы, лишь некоторые поднимаются в кроны саксаула для охоты или сбора пади.
Встает вопрос: каким же образом такое число видов может существовать в одном ярусе биогеоценоза. Прежде всего представители двух имеющихся здесь биоморф резко отличаются по трофическим связям и занимают разные трофические уровни. Естественно, что пищевая конкуренция между этими группами практически отсутствует. Более детальное изучение питания зоофагов показало, что в пределах этой биоморфы можно выделить три трофические группы, у которых питание настолько различно, что конкуренция между ними практически исключена. Прежде всего следует отметить крайне специализированное питание Crematogaster subdentata — почти исключительно тлями. Большие различия в составе пищи наблюдаются у герпетобионтов-зоофагов, ведущих дневной и ночной образ жизни. Ночью в пустыне на поверхности почвы появляются медлительные беспозвоночные, такие, как термиты, происходит линька саранчовых. Днем же активны лишь подвижные или покрытые плотными покровами насекомые, гораздо менее доступные как добыча. Поэтому, если ночные муравьи являются истинными зоофагами, дневные питаются по преимуществу трупами и лишь самые крупные и подвижные из них ( Catagtyphis setipes, C. foreli) добывают значительное количество живых насекомых, таких, как цикадки, саранчовые, имаго жуков и клопов, гусеницы. Единственный вид пищи, поедаемой в больших количествах представителями обеих групп, — термиты, но конкуренция из-за этого вида пищи не может быть значительной, так как, во-первых, муравьи поедают лишь незначительную часть биомассы термитов (Захаров, 1975а) и, во-вторых, Catagtyphis ловят этих насекомых главным образом во время лёта (т. е. уничтожают крылатых половых особей и сторожей), a Camponotus — во время постройки галерей и фуражировки (т. е. строителей и фуражиров). Различия в питании наблюдаются и у крупных карпофагов (муравьев-жнецов). У части видов (группа M. excursionis) фуражиры действуют на территории как одиночные сборщики и собирают рассеянные семена. Обычно эти муравьи активны в пустынях Средней Азии ночью. Большинство же видов Messor организуют групповую фуражировку и собирают урожай на участках с только что созревшими семенами эфемеров.
В пределах одной трофической группы у муравьев разных размеров различаются и размерные категории пищи. Так, Tetramorium schneideri (средний сухой вес рабочего p=0,5 мг) заготавливает семена весом 0,10—0,35 мг (средний вес 0,24 мг), a Messor variabilis (p=5,3 мг) — 1,8—13,7 мг (средний вес 6,3 мг). Более детальная картина дифференциации питания карпофагов, обитающих на одной территории, по разным размерным категориям пищи будет показана ниже. Аналогичная зависимость наблюдается и при сравнении питания зоофагов. Средний вес доставляемой в гнездо добычи у Acantholepis semenovi (p=0,23 мг) равен 0,79±0,06 (n=57), у Cataglyphis pallida (p=0,9 мг) — 4,95±0,67 мг (n=46), а у Cataglyphis setipes (p=6,7 мг) — 11,45± 1,09 мг (n=167) (рис:3).
Несколько модифицируя предложенную нами ранее (Длусский, 1975а) систему, можно предложить следующую классификацию биоморф и трофических групп муравьев:
A. Геобионты
A3 — зоофаги
АА — афидофаги
АК — клептобионты
Б. Стратобионты
БЗ — зоофаги
ВЗ — дневные зоонекрофаги и зоофаги
ВН — ночные зоофаги
ВК — карпофаги и карпонекрофаги
ВКо — одиночные сборщики семян
ВКг — виды, собирающие семена группами
ВМ — мицетофаги (в Палеарктике не встречаются)
Г. Хортобионты
ГЗ — зоофаги и зоонекрофаги
Д. Дендробионты
ДА — афидофаги
ДЗ — зоофаги
Кроме того, в пределах каждой трофической группы мы выделяем шесть размерных классов, взяв в качестве показателя размеров средний сухой вес рабочего:
1 — менее 0,1 мг
2 — 0,1—0,25 мг
3 — 0,25—1,0 мг
4 — 1,0—2,5 мг
5 — 2,5—10 мг
6 — более 10 мг
В дальнейшем тексте используется буквенно-цифровой индекс трофических групп, например: ВК-3 — герпетобионт-карпофаг 3-го размерного класса.
Не случайно в самостоятельную трофическую группу не были выделены трофобионты-зоофаги, или собиратели жидкой пищи, как это делают многие авторы (Арнольди, 1937, 1968; Culver, 1972; Whitford, 1978а). Это имело бы смысл при оценке роли муравьев в биогеоценозе, но, как мне кажется, не имеет значения при изучении структуры коадаптивных комплексов и изучении коадаптивной эволюции пустынных муравьев. Во-первых, в настоящей пустыне облигатные трофобионты существовать не могут, так как тли, выделяющие падь, встречаются лишь в немногих пустынных сообществах (на саксауле, кандыме и некоторых растениях солончаков) и лишь короткое время. Во-вторых, конкуренция из-за пади между трофобионтами практически исключена, так как муравьи могут создавать любое нужное количество колоний тлей. Возможно, существует конкуренция между дневными зоонекрофагами за эксудаты пустынных растений, но вопрос этот никем не изучался.
У видов, входящих в один размерный класс одной трофической группы, в одинаковых условиях состав пищи почти идентичен. Состав пищи видов, находящихся в соседних размерных классах, несколько различается, при разнице в два класса лишь частично перекрывается, а при разнице в три класса практически различен. Так, например, у Cataglyphis setipes (ВЗ-5) лишь 3,2% (по весу) составляют объекты питания, общие с Cataglyphis pallida (ВЗ-3), и 0,13% — общие с Acantholepis semenovi (ВЗ-2). В питании C. pallida пищевые объекты, общие с A. semenovi, составляют уже 26%. Хотя весь спектр добычи представителей низших размерных классов входит в спектр добычи более крупных муравьев той же трофической группы (например, все насекомые, найденные в питании C. pallida и A. semenovi, найдены и у C. setipes), конкуренция при добывании этих видов пищи не должна быть слишком сильной, так как работа крупных фуражиров при добывании слишком мелких пищевых объектов может оказаться неэффективной. Действительно, если сбор трупов муравьев-жнецов фуражирами C. setipes целесообразен, на что указывает целый ряд особенностей поведения этих муравьев, то энергетические затраты на поиск трупов Monomorium или Plagiolepis вряд ли окупаются. Реально единственными кормовыми объектами, из-за которых конкурируют C. setipes и C. pallida, являются трупы муравьев-жнецов (свежий вес 4—5 мг), а C. pallida и A. semenovi — трупы муравьев весом около 1 мг ( Tetramorium schneideri, Crematogaster subdentata). В дальнейшем мы будем условно принимать, что виды, входящие в один размерный класс одной трофической группы одной биоморфы, занимают одну трофическую нишу.
В группе герпетобионтов карпофагов (ВК) резко доминируют виды, • относящиеся к 2-му и 5-му размерным классам. Конкуренция между ними практически исключена. T. schneideri (ВК-3), по-видимому, в какой-то степени конкурирует с муравьями-жнецами (ВК-5). Во всяком случае, биомасса этого вида достигает максимума именно в тех типах пустынь, где биомасса муравьев-жнецов низка. Например, в саксаульниках Акыбая сухая биомасса Messor составляет в среднем 316 г/га и T. schneideri — 61 г/га, а в саксаульниках Джарда — соответственно 130 и 483 г/га. Для этой группы характерно полное отсутствие представителей 4-го размерного класса, что, несомненно, связано с высокой вариабельностью рабочей касты муравьев-жнецов.
В группе дневных зоонекрофагов (ВЗ) довольно равномерно представлены все размерные классы и число видов в каждом классе сравнительно невелико. Ночные герпетобионты-зоофаги (ВН) четко разбиваются на два размерных класса: с одной стороны, это группа из трех видов, имеющих близкие размеры (p=46 мг), и с другой — Camponotus xerxes, самый крупный из наших муравьев (p=7—55 мг).
При анализе количественного распределения муравьев в различных литоэдафических типах пустынь (табл.4) и разных растительных ассоциациях песчаных пустынь (табл.5) Средней Азии бросается в глаза общая закономерность: в каждом конкретном местообитании с однородными условиями встречается несколько (чаще всего два) видов мелких карпофагов (ВК-2), один —три вида муравьев-жнецов (ВК-5), чаще всего один ВКо и один ВКг, и, как правило, не более чем по одному обычному виду остальных размерных классов каждой трофической группы. Единственное исключение объясняется неравномерностью условий: в эфемерово-полынной песчаной пустыне в горах Кульджуктау чередуются участки плотных загипсованных песков, где обитает Cataglyphis emeryi, и наносных песков, где обитает C. pallida (оба ВЗ-3). Общая биомасса и соотношение биомасс представителей разных трофических групп (табл.6), число видов муравьев и число заполненных трофических ниш каждого конкретного местообитания определяются общей продуктивностью, динамикой продукции и флористическим составом биогеоценозов.
Здесь сразу следует оговориться, что до 1970 г. мы использовали главным образом маршрутный метод учета численности муравьев (Длусский, 1965б). При целом ряде преимуществ (возможность сочетать учеты со сборами фауны, охват большой территории) этот метод является лишь относительным: при нем несколько завышается численность крупных Cataglyphis и большинства видов Messor и сильно занижается численность мелких муравьев. Однако, поскольку все учеты были сделаны одним исследователем (автором), сравнение разных участков вполне возможно и отражает общую картину. Одновременно в некоторых случаях делался учет мелких муравьев на площадках 10x10м. В 1970 г. в Акыбае нам потребовалось сделать абсолютный учет численности. Были использованы площадки 20X20 м для учета Messor и Cataglyphis и площадки 5x5 для учета остальных муравьев. Для быстрого выявления всех гнезд на малых площадках за 2—3 ч до учета выкладывались кормушки с манной крупой и сахарным сиропом. Полученные данные о числе гнезд пересчитывались на условную биомассу (среднее число рабочих в семье, округленное до 0,5 порядка, X средний сухой вес рабочего). Население семьи определялось путем раскопки гнезд. Подобным же образом удалось пересчитать часть данных предыдущих лет. В 1978 г. в Варганзе мы применили ту же методику учетов.
В наиболее наглядном виде типичная структура «пустынных» коадаптивных комплексов муравьев представлена на пухлых солончаках (табл.4). Так, на сазовом солончаке в предгорьях Кульджуктау (оазис Аяк-Гужумды) при довольно высокой общей численности и биомассе (659 г/га) обитают всего пять видов муравьев, резко отличающихся по спектрам питания: два, отличающихся размерами карпонекрофага, — Cardiocondyla uljanini (ВК-1, p=0,07 мг) и Monomorium kusnezovi (ВК-2, p=0,24 мг), мелкий некрофаг Tapinoma karavaievi (ВЗ-2, p=0,44 мг), крупный дневной зоонекрофаг ![]() Cataglyphis aenescens (ВЗ-4, p=1,80 мг) и ночной зоофаг Camponotus turcestanicus (ВН-5, p=6,36 мг). Резкое доминирование зоонекрофагов (табл.6) объясняется тем, что на солончаках Кызылкумов обитает большое количество насекомых, связанных с тамариксом, семена которого муравьями не поедаются, и продукция пригодной для муравьев животной пищи больше, чем растительной.
Cataglyphis aenescens (ВЗ-4, p=1,80 мг) и ночной зоофаг Camponotus turcestanicus (ВН-5, p=6,36 мг). Резкое доминирование зоонекрофагов (табл.6) объясняется тем, что на солончаках Кызылкумов обитает большое количество насекомых, связанных с тамариксом, семена которого муравьями не поедаются, и продукция пригодной для муравьев животной пищи больше, чем растительной.
По общей численности и богатству фауны муравьев настоящие (ксерофитные) пустыни можно расположить в следующий ряд: бедлендовые, такыры, барханы, гипсовые, каменистые, галечниково-песчаные (солонцеватые), глинистые лессовые, эфемерово-полынные песчаные, песчаные с саксауловыми лесами. В бедлендовой пустыне близ оазиса Аяк-Гужумды муравьи вообще не были обнаружены. Биомасса муравьев на такырах вблизи пос. Артезианский была 0,7 г/га, а на подвижных барханах в Каррыкуле 24 г/га. Муравьи представлены здесь в основном дневными некрофагами, питающимися трупами случайно залетевших или занесенных ветром насекомых. На барханах в окрестностях Иолотани (Юго-Восточные Каракумы) наряду с некрофагом Cataglyphis pallida изредка встречается карпофаг Messor excursionis, а в Репетеке — Monomorium barbatulum. Эти муравьи, по-видимому, питаются семенами, занесенными ветром.
В полынно-солянковых (ass, Artemisia turanica + Salsola rigida + S. arbuscula) и тасбиюргуновых (ass. Nanophyton crinaceum) гипсовых пустынях Кызылкумов (табл.4, табл.6) общая биомасса муравьев равна 140 г/га. Фон здесь составляют всего шесть видов, занимающих пять различных трофических ниш: Tetramorium inerme (ВК-2), Monomorium barbatulum (ВК-2), Tetramorium schneideri (BK-3), Messor variabilis (BKo-5), Plagiolepis pallescens (BK-1) и Cataglyphis emeryi (B3-3). T. inerme и P. pallescens живут в кустиках полыни и практически не выходят за их пределы. M. barbatulum, T. schneideri и M. variabilis, резко отличающиеся размерами (р равно соответственно 0,14; 0,48 и 5,34 мг), собирают на открытых местах семена эфемеров, а C. emeryi — трупы насекомых. Скудность эфемеровой растительности приводит к тому, что биомасса карпофагов невелика и ненамного превышает биомассу некрофагов. Сходную структуру имеют комплексы муравьев полынных ассоциаций (ass. Artemisia turanica) каменистой пустыни в останцовых горах Кульджуктау (табл.4). Здесь встречаются семь обычных видов, равномерно заполняющих семь трофических ниш (ВК-1, ВК-2, ВК-3, Вкг-5, ВЗ-1, ВЗ-3, ВЗ-4), причем Tetramorium inerme (ВК-2) и Plagiolepis pallescens (ВЗ-1), как и в гипсовой пустыне, обитают и добывают пищу в кустиках полыни.
Для галечниково-песчаной солонцеватой пустыни (ass. Salsola gemmascens) в древнем русле Зеравшана (Дарья-сай, табл.4) и эфемерово-полынных (ass. Artemisia turanica + ephemeretum) песчаных пустынь (табл.5) Кызылкумов характерно резкое преобладание зоофагов и зоонекрофагов. Численность и видовое разнообразие карпофагов и мелких некрофагов ( P. pallescens) здесь существенно не отличаются от гипсовых и каменистых пустынь. В то же время численность дневных некрозоофагов среднего размера — Cataglyphis emeryi (на загипсованных песках) и C. pallida — здесь на порядок выше. Видимо, в связи с тем, что именно в этих типах пустынь в Кызылкумах наблюдается наибольшее количество термита Anacanthotermes ahngerianus, здесь появляются крупный дневной зоофаг Cataglyphis foreli (ВЗ-5) и ночные зоофаги Camponotus turcestanus и C. fedtschenkoi (ВН-5). Прямыми наблюдениями в Дарья-сае было установлено, что именно термиты являются основным компонентом пищи C. foreli. Питание C. turcestanus и C. fedtschenkoi не изучалось, но, судя по аналогии с другими видами рода, эти муравьи также должны в основном поедать термитов. В целом в каждом из этих типов пустынь обычны 9—10 видов, занимающих 7—8 трофических ниш.
Для фауны эфемеровой лессовой пустыни в окрестностях Мубарека (табл.4, табл.6) характерно при не особенно большой общей биомассе муравьев (387 г/га) резкое преобладание карпофагов над зоонекрофагами (биомассы соответственно 361 и 26 г/га). Основная продукция фитомассы создается здесь эфемеровой растительностью и летом глинистая лессовая пустыня представляет собой безжизненную равнину. В таких условиях вполне могут существовать карпофаги, создающие весной запасы пищи (семян) в гнездах. Для зоонекрофагов, не создающих значительных запасов пищи и вынужденных добывать пищу летом, эти условия оказываются крайне неблагоприятными, и поэтому биомасса их в лессовой пустыне приблизительно равна биомассе муравьев на подвижных барханах. Зоонекрофаги представлены в лессовой пустыне тремя видами, резко различающимися по размерам: Acantholepis semenovi (ВЗ-2, p=0,23 мг), Cataglyphis cinnamomea (ВЗ-4, p=1,43 мг) и C. foreli (ВЗ-5, p=7,45 мг). Карпофаги и карпонекрофаги представлены Monomorium kusnezovi (ВК-2), Tetramorium inerme (ВК-2), T. schneideri (ВК-3), Messor aralocaspius (ВКг-5) и M. vicinus (ВКг-5). Стации местообитания M. kusnezovi и T. inerme несколько отличаются: первый вид предпочитает понижения, а второй — участки с наносами песка.
Наибольшим богатством видов и наибольшей сложностью структуры коадаптивных комплексов отличаются песчаные пустыни с саксауловыми лесами (табл.5, табл.6), особенно расположенные в районах с неглубоким (3—5 м) залеганием грунтовых вод (Акыбай, Шафиркан). Характерными особенностями мирмекофауны таких пустынь является, во-первых, полное использование трофических ниш, и, во-вторых, большое количество видов мелких карпофагов (ВК-2), обитающих совместно. В качестве примера можно разобрать хорошо изученный (Длусский, 1975а) приспевающий саксаульник на песчаной почве в Акыбайском лесничестве Марыйского лесхоза ТССР. Общая биомасса муравьев в этом саксаульнике— 1827 г/га, причем биомасса карпофагов (801 г/га) составляет около 44%. Здесь обитают 16 обычных видов, занимающих 10 трофических ниш. Группа мелких карпофагов (ВК-2 /представлена пятью видами ( ![]() Monomorium gracillimum, M. barbatulum, M. kusnezovi, Tetramorium inerme, Pheidole arenarum), причем все эти виды могут встречаться под кроной одного дерева и входы в их гнезда представляют сложную мозаику (рис:4). Из других муравьев, питающихся семенами, здесь обычны Cardiocondyla uljanini (ВК-1), Tetramorium schneideri (ВК-3) и три вида муравьев-жнецов — Messor aralocaspius, M. intermedius (ВК-5) и M. lamellicornis (ВКо-5). Дневные зоонекрофаги представлены Plagiolepis pallescens (ВЗ-1), Acantholepis semenovi (ВЗ-2), Cataglyphis pallida (ВЗ-3) и C. setipes (ВЗ-5), ночные зоофаги — Camponotus xerxes (ВН-6).
Monomorium gracillimum, M. barbatulum, M. kusnezovi, Tetramorium inerme, Pheidole arenarum), причем все эти виды могут встречаться под кроной одного дерева и входы в их гнезда представляют сложную мозаику (рис:4). Из других муравьев, питающихся семенами, здесь обычны Cardiocondyla uljanini (ВК-1), Tetramorium schneideri (ВК-3) и три вида муравьев-жнецов — Messor aralocaspius, M. intermedius (ВК-5) и M. lamellicornis (ВКо-5). Дневные зоонекрофаги представлены Plagiolepis pallescens (ВЗ-1), Acantholepis semenovi (ВЗ-2), Cataglyphis pallida (ВЗ-3) и C. setipes (ВЗ-5), ночные зоофаги — Camponotus xerxes (ВН-6).
Наличие подстилки, которая образует под кронами саксаулов слой толщиной в несколько сантиметров, естественно, сказывается на структуре многовидовых ассоциаций муравьев. Если сравнить плотность входов в гнезда на подстилке под кронами и на открытых участках, сразу обнаруживается различие между видами. У ![]() M. gracillimum в подстилке открываются 0,45 вх/м2, а в песке между саксаулами 0,14 вх/м2, у M. barbatulum — соответственно 0,12 и 0,22 вх/м2, у M. kusnezovi 0,07 и 0,16 вх/м2, у T. inerme 0,10 и 0,02 вх/м2, у Ph. arenarum, P. pallescens и C. uljanini только в подстилке (0,07; 0,06 и 0,05 вх/м2). 2 мая 1971 г. на пробных площадках, изображенных на рис:4, было одновременно выставлено в шахматном порядке (сторона квадрата 1 м) 251 кормушка с манной крупой, которая охотно поедается мелкими муравьями-карпофагами. 74 кормушки были расположены на подстилке под кронами саксаулов, 29 — на границе подстилки и песчаных участков (полоса шириной 0,5 м) и 148 — на открытых песчаных участках между саксаулами.
M. gracillimum в подстилке открываются 0,45 вх/м2, а в песке между саксаулами 0,14 вх/м2, у M. barbatulum — соответственно 0,12 и 0,22 вх/м2, у M. kusnezovi 0,07 и 0,16 вх/м2, у T. inerme 0,10 и 0,02 вх/м2, у Ph. arenarum, P. pallescens и C. uljanini только в подстилке (0,07; 0,06 и 0,05 вх/м2). 2 мая 1971 г. на пробных площадках, изображенных на рис:4, было одновременно выставлено в шахматном порядке (сторона квадрата 1 м) 251 кормушка с манной крупой, которая охотно поедается мелкими муравьями-карпофагами. 74 кормушки были расположены на подстилке под кронами саксаулов, 29 — на границе подстилки и песчаных участков (полоса шириной 0,5 м) и 148 — на открытых песчаных участках между саксаулами. ![]() M. gracillimum посещал 66% кормушек на подстилке, 29% на границе и 26% на песке, тогда как M. barbatulum — 8,32% на подстилке и 38% на песке. T. inerme посещал главным образом кормушки, расположенные на границе микростаций (4% на подстилке, 17% на границе и 0,7% кормушек на песке), а T. schneideri — только вне подстилки (0,3% на подстилке и 7% кормушек на песке). Явственного предпочтения в выборе зоны фуражировки у Ph. arenarum, M. kusnezovi и C. uljanini обнаружить не удалось.
M. gracillimum посещал 66% кормушек на подстилке, 29% на границе и 26% на песке, тогда как M. barbatulum — 8,32% на подстилке и 38% на песке. T. inerme посещал главным образом кормушки, расположенные на границе микростаций (4% на подстилке, 17% на границе и 0,7% кормушек на песке), а T. schneideri — только вне подстилки (0,3% на подстилке и 7% кормушек на песке). Явственного предпочтения в выборе зоны фуражировки у Ph. arenarum, M. kusnezovi и C. uljanini обнаружить не удалось.
Таким образом, все виды мелких муравьев-карпофагов, обитающих в саксаульнике, можно разделить на две группы: виды, живущие и собирающие пищу преимущественно в подстилке под саксаулами, и виды, живущие и собирающие пищу преимущественно на открытых участках. Открытые участки эксплуатируют главным образом три вида мелких карпофагов, средние веса рабочих которых соотносятся приблизительно как 1:2:4 ( M. barbatulum, p=0,14; M. kusnezovi, p=0,24; T. schneideri, p=0,48 мг). Конкуренция между этими тремя видами ослабляется также и потому, что M. kusnezovi предпочитает более влажные участки с однолетними солянками, а два других — более сухие участки с эфемерами. В подстилке обитают три вида карпофагов, рабочие которых имеют приблизительно одинаковые размеры ( C. uljanini, p=0,07; T. inerme, p=0,11; Ph. arenarum, p=0,12 мг), и ![]() M. gracillimum, средние размеры рабочих которого значительно больше (p=0,27 мг), но размеры рабочих (а следовательно, и размер заготавливаемой добычи) сильно варьируют. Поскольку под кронами саксаулов эфемеры встречаются реже, чем на открытых участках, можно предположить, что в значительной степени эти виды (или часть из них) переключаются на зоонекрофагию в слое подстилки, т. е. становятся стратобионтами-зоонекрофагами, и вступают в конкуренцию с обитающим здесь же P. pallescens (p=0,08 мг). Каковы взаимоотношения между всеми этими видами — пока неясно. Заметны лишь некоторые тенденции. Так, C. uljanini и T. inerme обитают и собирают пищу преимущественно по краю пятна подстилки. Входы в гнезда Ph. arenarum и P. pallescens открываются, как правило, вблизи основания стволов саксаула. Рабочие P. pallescens ходят чаще всего по стволам, почти не заходя на тонкие ветви и постоянно заглядывая под отслаивающиеся чешуйки коры, a Ph. arenarum на деревьях отмечались редко. Фуражиры
M. gracillimum, средние размеры рабочих которого значительно больше (p=0,27 мг), но размеры рабочих (а следовательно, и размер заготавливаемой добычи) сильно варьируют. Поскольку под кронами саксаулов эфемеры встречаются реже, чем на открытых участках, можно предположить, что в значительной степени эти виды (или часть из них) переключаются на зоонекрофагию в слое подстилки, т. е. становятся стратобионтами-зоонекрофагами, и вступают в конкуренцию с обитающим здесь же P. pallescens (p=0,08 мг). Каковы взаимоотношения между всеми этими видами — пока неясно. Заметны лишь некоторые тенденции. Так, C. uljanini и T. inerme обитают и собирают пищу преимущественно по краю пятна подстилки. Входы в гнезда Ph. arenarum и P. pallescens открываются, как правило, вблизи основания стволов саксаула. Рабочие P. pallescens ходят чаще всего по стволам, почти не заходя на тонкие ветви и постоянно заглядывая под отслаивающиеся чешуйки коры, a Ph. arenarum на деревьях отмечались редко. Фуражиры ![]() M. gracillimum (и входы в гнезда) равномерно распределены по всему пятну подстилки и выходят на открытые участки вблизи крон.
M. gracillimum (и входы в гнезда) равномерно распределены по всему пятну подстилки и выходят на открытые участки вблизи крон.
Приблизительно так же выглядит и структура коадаптивных комплексов молодых саксаульников на песчаной почве и приспевающих саксаульников на плотной почве в том же районе (Длусский, 1975а) и спелых саксаульников на песчаной почве на юго-западной границе Кызылкумов (Шафиркан, табл.5). В молодых саксаульниках на песчаной почве видовой состав муравьев примерно тот же, что и в приспевающем, но среди мелких карпофагов на первое место по численности и биомассе выходит Ph. arenarum, общая биомасса муравьев меньше (1446 г/га), а удельный вес карпофагов выше (746 г/га, или 52%), чем в приспевающих. Еще меньше биомасса муравьев в саксаульниках на плотной почве (548 г/га). Биомасса карпофагов здесь относительно выше, чем в саксаульниках на песках (365 г/га, или 67%). M. lamellicornis замещается M. variabilis (ВКо-5), а C. pallida — C. emeryi (ВЗ-3). Комплексы муравьев в песчаной пустыне с кандымом и песчаной акацией в глубинных районах Кызылкумов (Кульджуктау, табл.5) и в старых естественных саксаульниках Центральных Каракумов (Джард, табл.5, табл.6) занимают промежуточное положение между комплексами эфемерово-полынных песчаных пустынь и богатых саксаульников Шафиркана и Акыбая.
Наибольшую сложность представляет мирмекофауна комплексных типов пустынь, таких, как грядово-такырные комплексы, где узкие полосы такыров чередуются с грядами задернованных и барханных песков (Длусский, 19746), ячеистые пески с «блюдцами» глинистой почвы, часто засоленной, или саи (долины, промытые дождевыми потоками) в предгорьях останцовых гор с участками сазовых солончаков в местах выхода соленых грунтовых вод (табл.7). Число (но не общая биомасса) видов в таких биогеоценозах непропорционально высоко в сравнении с их продуктивностью. Так, например, в грядово-такырном комплексе Центральных Каракумов (Каррыкуль) встречается 16 обычных видов, а в не менее продуктивных саксауловых лесах Центральных Каракумов (Джард) — только 10. В саях с выходом грунтовых вод в предгорьях Кульджуктау найдено 15 обычных видов, а в расположенных в том же районе более продуктивных солончаках и эфемерово-полынных пустынях — 5 и 8.
Такое разнообразие объясняется тем, что многие муравьи предпочитают селиться на границах растительных ассоциаций, поскольку оптимальные условия для постройки гнезд могут не совпадать с оптимальными условиями для фуражировки. Кроме того, одновременное использование двух различных растительных ассоциаций обеспечивает более разнообразный состав кормов и, следовательно, более стабильную кормовую базу. В качестве примера можно рассмотреть распределение муравьев по профилю грядово-такырного комплекса Центральных Каракумов (рис:5). На схеме видно, что гнезда муравьев-жнецов (ВК-5) и C. xerxes (ВН-6) располагаются преимущественно на такырах, где плотная почва и близкое залегание грунтовых вод создают благоприятные условия для постройки гнезд (Длусский, 1975б), но по большей части вблизи границы такыров и песчаных бугров. Наблюдения показали, что фуражировка этих муравьев происходит в основном на более богатых растительностью песчаных грядах. C. foreli (ВЗ-5), гнездящийся на песчаных грядах, напротив, иногда фуражирует на такырах. В то же время более мелкие виды, занимающие сходные трофические ниши, например дневные некрозоофаги Cataglyphis pallida (p=0,89 мг) и C. cinnamomea (p=1,43 мг), добывают пищу в тех же ассоциациях, где строят гнезда.
Подводя итоги, можно сказать, что структура коадаптивных комплексов во всех пустынях Казахстана и Средней Азии в общих чертах сходна и в то же время принципиально отличается от структуры комплексов гумидных областей. Подавляющее большинство входящих в них видов относятся к герпетобионтам и добывают пищу на поверхности почвы. Однако узкая пищевая специализация обеспечивает возможность сосуществования в одном биогеоценозе достаточно большого числа видов, не меньшего, чем в гумидных биогеоценозах той же климатической зоны. Дифференциация по питанию идет несколькими путями: по типу пищи (карпофаги, дневные зоонекрофаги, ночные зоофаги, афидофаги, клептобионты), по размеру пищи, довольно тесно скоррелированному с размерами муравьев-фуражиров и, для мелких муравьев, по зонам, в которых собирается пища (виды, собирающие пищу внутри кустиков полыни и на открытых участках с эфемерами). В биогеоценозе без древесной растительности муравьи могут заполнять до 13 трофических ниш: клептобионты (один вид), мелкие карпофаги, обитающие внутри кустиков полыни (один вид), карпофаги, собирающие пищу на открытых участках (четыре размерных класса), мелкие некрофаги, обитающие внутри кустиков полыни (один вид), дневные некрозоофаги и зоонекрофаги (четыре размерных класса), ночные зоофаги (два размерных класса). При наличии древесной растительности (саксаульники) количество возможных трофических ниш увеличивается: появляются муравьи-дендробионты (два вида с разными спектрами питания) и формируется комплекс видов, связанных с подстилкой.
В каждом конкретном биогеоценозе встречается, как правило, не более одного вида, занимающего одну трофическую нишу. Исключение составляют рассмотренные выше комплексы видов, обитающих в подстилке под саксаулами, и муравьи-жнецы. У последних возможность сосуществования разных видов в одном биогеоценозе объясняется сложными пространственно-временными взаимоотношениями, которые будут подробно рассмотрены ниже. Однако ни в одном из изученных биогеоценозов все потенциально возможные трофические ниши не оказываются заполненными. Количество заполненных ниш, общее обилие муравьев и соотношение биомасс представителей разных трофических групп определяется общей продуктивностью, соотношением продукции доступных для муравьев семян и мелких животных и характером динамики продукции фитомассы каждого конкретного биогеоценоза.
Какие виды будут занимать ту или иную трофическую нишу в каждом конкретном биогеоценозе, определяется географической широтой местности и литоэдафическим типом пустыни. По мере продвижения на север общее число видов постепенно уменьшается, а компенсации почти не происходит: в пустынях Казахстана появляются Proformica epinotalis, замещающая Acantholepis semenovi пустынь Средней Азии, дендробионт-зоофаг Camponotus semirufus, не имеющий аналогов в пустынях Средней Азии, и несколько видов муравьев-жнецов. В том случае, когда размерный класс трофической группы представлен одним видом, для этого вида характерна высокая эвритопность. Так, Tetramorium schneideri и Plagiolepis pallescens были обнаружены нами во всех изученных биотопах, кроме такыров, солончаков и подвижных барханов. Такой же эвритопностью отличаются Camponotus xerxes и Acantholepis semenovi в пределах своих ареалов. В то же время, если в одном размерном классе зоофагов находится несколько видов, они занимают разные стации. Так, Cataglyphis setipes связан преимущественно с саксаульниками и глинистыми пустынями, а C. foreli — с пустынями без древесной растительности с песчаной или супесчаной почвой; вместе эти виды I практически не встречаются. Среди зоофагов средних размеров Cataglyphis pallida обитает на легких песчаных почвах, C. emeryi — на супесчаных и суглинистых, C. cinnamomea — на глинистых и каменистых, ![]() C. aenescens — в оазисах и на солончаках в пустынях Средней Азии или в глинистых и суглинистых пустынях — в тех районах, где отсутствует C. cinnamomea. В группе ночных зоофагов закономерности распределения Camponotus turcestanus и C. fedtschenkoi пока неясны, но эти виды также обычно не встречаются вместе. C. turcestanicus обитает на солончаках и в оазисах. Аналогичная картина наблюдается и у муравьев-жнецов из группы M. excursionis: M. excursionis встречается на легких песчаных почвах, M. lamellicornis — на более плотных супесчаных почвах и в саксаульниках, M. variabilis — на глинистых и суглинистых почвах.
C. aenescens — в оазисах и на солончаках в пустынях Средней Азии или в глинистых и суглинистых пустынях — в тех районах, где отсутствует C. cinnamomea. В группе ночных зоофагов закономерности распределения Camponotus turcestanus и C. fedtschenkoi пока неясны, но эти виды также обычно не встречаются вместе. C. turcestanicus обитает на солончаках и в оазисах. Аналогичная картина наблюдается и у муравьев-жнецов из группы M. excursionis: M. excursionis встречается на легких песчаных почвах, M. lamellicornis — на более плотных супесчаных почвах и в саксаульниках, M. variabilis — на глинистых и суглинистых почвах.
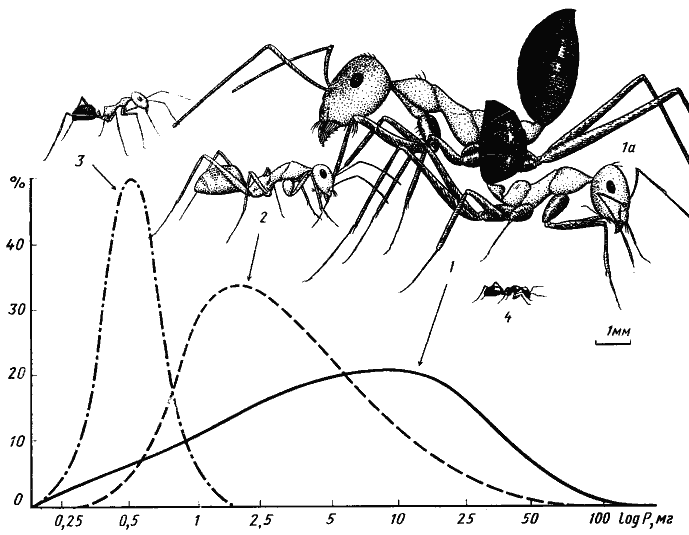
Рис. 3. Сравнительные размеры четырех основных видов дневных зоонекрофагов песчаной пустыни (Акыбай) и частотное распределение (в %) добычи трех видов по весовым классам (усредненные данные)
1, 1а — средний и крупный рабочие Cataglyphis setipes;
2 — C. pallida;
3 — Acantholepis semenovi;
4 — Plagiolepis pallescens
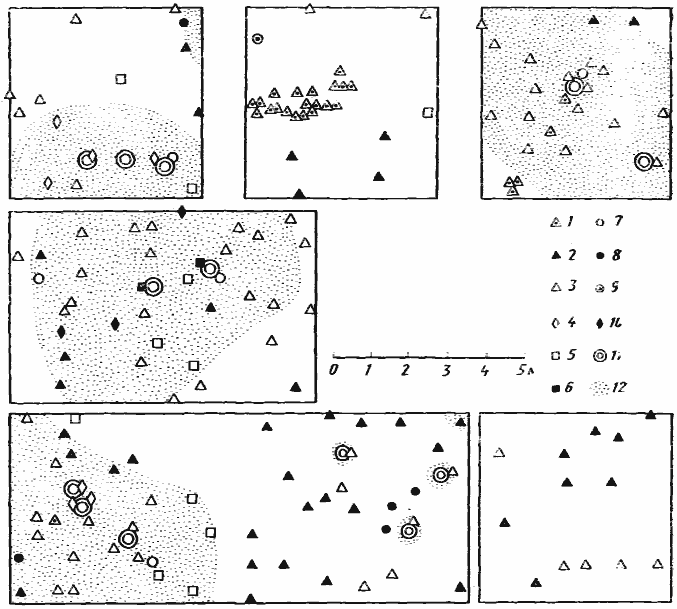
Рис. 4. Размещение входов в гнезда муравьев на шести пробных площадках в саксауловом лесу
Акыбай, 2 мая 1971 г.)
1 — Monomorium kusnezovi;
2 — M. barbatulum;
3 —
4 — Pheidole pallidula;
5 — Tetramorium inerme;
6 — Crematogaster subdentata;
7 — Plagiolepis pallescens;
8 — Acantholepis semenovi;
9 — Cataglyphis pallida;
10 — Cardiocondyla uljanini;
11 — саксаулы;
12 — подстилка под саксаулами
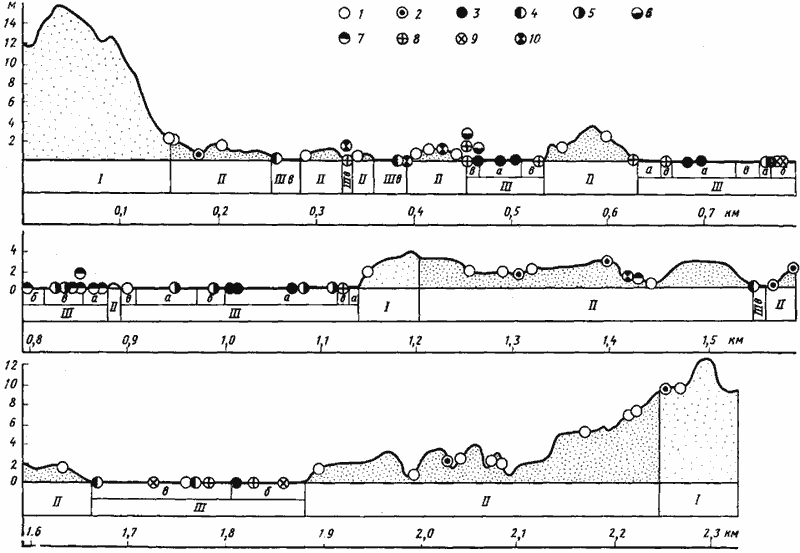
Рис. 5. Распределение муравьев по профилю грядово-такырного комплекса в Центральных Каракумах
(Каррыкуль) [Длусский, 19746]
I — незакрепленные пески; II — закрепленные пески (общая проективная полнота 30—45%); III — такыровидные поверхности: а — лишенные высшей растительности; б — без песчаной присыпки с полынным тетырником с проективной полнотой 10—15%; в — с песчаной присыпкой и проективной полнотой растительности 20—25%
1 — Cataglyphis pallida; 2 — C. foreli; 3 — C. cinnamomea; 4 — Messor intermedins; 5 — M. variabilis; 6 — M. aralocaspius; 7 — Pheidole arenarum; 8 — Camponotus xerxes; 9 — C. turkestanus; 10 — Acantholepis semenovi
организм, обитающий в древесном ярусе растительности.
отряд общественных насекомых. Общины, разделенные на касты, состоят из крылатых и бескрылых особей. Строят подземные и наземные (до 15 м высотой) гнезда (термитники). Ок. 2600 видов, главным образом в тропиках; в России 2 вида: один в районе Сочи, второй, вероятно, завезенный из Японии или Китая, во Владивостоке. Многоядны. Разрушают древесину, портят бумагу, кожу, сельскохозяйственные продукты и др. Термиты, обитающие в почве, полезны как почвообразователи.
организм, обитающий среди растительных или иных органических остатков, на поверхности почвы.
организм, питающийся растениями
организм, питающийся исключительно другими животными
сладкая густая жидкость, выделяемая тлями, червецами и другими насекомыми, питающимися соком растений. Медоносная пчела собирает падь с листьев и перерабатывает в падевый мед.
эволюционно сложившаяся, пространственно ограниченная, длительно самоподдерживающаяся, однородная экологическая система, в которой функционально взаимосвязаны живые организмы и окружающая их абиотическая среда.
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
животные, не имеющие позвоночника: простейшие, губки, кишечнополостные, черви, моллюски, членистоногие, иглокожие и др.; примерно 3/4 видов всех животных.
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
ксеробионт
взрослая особь
общая масса особей одного вида, группы видов или сообщества в целом на единицу поверхности или объема местообитания. Биомасса растений называется фитомассой, биомасса животных зоомассой.
Всего 965 видов.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
организм, питающийся семенами растений.
Всего 106 видов.
Старые названия: Acantholepis semenovi, Acantholepis transcaspiensis
ксеробионт
организм, обитающий на поверхности и в толще пород суши.
организм, питающийся тлями (Aphididae) и их паразитоидами
организм, питающийся за счет воровства
организм, обитающий в лесной подстилке.
(зоонекрофаг), организм, питающийся мертвыми животными, гетеротроф
организм, питающийся мицелием грибков
организм, живущий в траве
взаимовыгодные отношения между муравьями и выделяющими падь насекомыми: слизывая падь, муравьи охраняют этих насекомых.
необратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.
итал. obligato, "обязательный", необходимый, существенный
Всего 60 видов.
у муравьев имеется три основные касты: самцы, самки и рабочие (бесплодные модифицированные самки).
ксеробионт
Старое название: Proformica deserta
Всего 65 видов.
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
галобионт
Subspecies: Monomorium indicum kusnezowi Santschi, 1928
гигрофил, галобионт
 Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунок
Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунокгигрофил, галобионт
Старое название: Cataglyphis flavigastra
олигогинны
Старое название: Cataglyphis cinnamomea
ксеробионт
 Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушитель
Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушительСтарое название: Monomorium gracillimum
Старое название: Pheidole arenarum Ruzsky
территориальная иерархическая группировка муравейников разных видов, обладающая устойчивой структурой, которая поддерживается поведенческими механизмами.
(гр. topos - место) - естественное жизненное пространство определенного биоценоза, совместно с которым он образует экосистему.






