






По-видимому, В. М. Вилер в работе о медовых муравьях (Wheeler, 1908а) впервые четко и полно изложил идею, которая затем повторялась во всех работах, посвященных как сравнению муравьев разных пустынь Земли, так и анализу фаун отдельных аридных областей (Wheeler, 1910; Forel, 1911; Emery, 1912а, 1920; Kusnezov, 1956; Bernard, 1958; Delye, 1968; и др.). Эту работу он заканчивает такими словами: «Большинство муравьев умеренных мезофитных районов имеет смешанную диету, состоящую из насекомых, пади и выделений растений. Когда такие виды переходят жить в пустыню или другие аридные районы, где длительная засуха летом и холод зимой заставляют растения и животных жить короткий сезон, они обычно избирают (take) одну из следующих адаптации:
1. Они могут усилить хищнические привычки, которыми они уже обладают, и стать неустрашимыми прожорливыми охотниками. Таким образом они могут собрать достаточное количество пищи даже в неблагоприятных условиях. Эта адаптация прекрасно выражена у Myrmecocystus Старого Света ( Cataglyphis. — Г. Д.), которые представлены огромным числом видов, подвидов и варьететов в пустынях Северной Африки. Ту же тенденцию проявляют, как оказалось, американские расы orblceps и mendax ( Myrmecocystus melliger, M. mendax. — Г. Д.).
2. Многие виды переходят к питанию и собиранию семян — очень распространенная адаптация в аридных областях, покрытых коротко живущими травянистыми растениями, показанная у видов Pogonomyrmex Нового Света, Messor (в Неоарктике — Veromessor. — Г. Д.), Solenopsis и Pheidole обоих полушарий и Holeomyrmex (= Monomorium. — Г.Д.), Oxyopomyrmex, Goniomma, Meranoplus, Pheidologeton Старого Света. Эти муравьи еще питаются насекомыми, когда их можно добыть, но семена представляют такой неистощимый и питательный вид пищи (food supply), что привычки к собиранию и запасанию семян в гнездах становятся высоко развитыми.
3. Многие виды медовых муравьев... начинают запасать сладкие выделения растений и экскременты тлей и кокцид в зобу физиологической касты «хранителей меда».
4. Некоторые муравьи могут жить и процветать в аридных районах, поскольку они культивируют и едят грибы. Этот образ жизни... наблюдается в единственной трибе американских ![]() Myrmicinae — Attini — и, видимо, зародился в буйных влажных тропических лесах. Затем несколько видов мигрировало в пустыни Северной Мексики и юго-западных Штатов, где они могут всегда найти растительные остатки для субстрата, на котором выращиваются грибы, и где эти нежные растения могут культивироваться на некоторой глубине, ниже сухой поверхности почвы» (Wheeler, 1908а, с. 395).
Myrmicinae — Attini — и, видимо, зародился в буйных влажных тропических лесах. Затем несколько видов мигрировало в пустыни Северной Мексики и юго-западных Штатов, где они могут всегда найти растительные остатки для субстрата, на котором выращиваются грибы, и где эти нежные растения могут культивироваться на некоторой глубине, ниже сухой поверхности почвы» (Wheeler, 1908а, с. 395).
Хотя эти утверждения получили широкое распространение и, несомненно, содержат рациональное зерно, полностью согласиться с ними трудно. Представление о том, что подвижные дневные муравьи ( Cataglyphis, некоторые Myrmecocystus, Dorymyrmex и т. д.) являются «неустрашимыми прожорливыми хищниками», явно основано на ложной аналогии с позвоночными-хищниками, берущими добычу «в угон». На самом деле это робкие пугливые насекомые. Если, например, выложить на участке, где совместно фуражируют ![]() C. aenescens и Messor denticulatus, труп крупного насекомого, вокруг него моментально собираются муравьи-жнецы. Бегунки обычно держатся поодаль, и время от времени один из них подбегает и пытается оторвать кусок, но моментально отскакивает в сторону, стоит другому муравью прикоснуться к нему антенной или даже резко повернуться. Если говорить об аналогиях с позвоночными, то Cataglyphis скорее можно уподобить классическим некрофагам. Для них характерны приспособления, обеспечивающие возможность контролировать большую территорию, на которой в разных местах случайна появляются источники пищи. На этом аналогии кончаются. Позвоночные обычно питаются трупами крупных животных, которые должны быть расчленены на месте. У них вырабатывается два пути адаптации: постоянное патрулирование территории с малыми затратами энергии (планирование грифов и марабу на большой высоте) или взаимное привлечение на больших расстояниях (характерный вой шакалов и гиен) (Lewic-Goodall, Lewic-Goodall, 1970). Муравьи-бегунки, напротив, обычно собирают трупы, которые могут быть принесены в гнездо одним фуражиром. Каждый фуражир обследует в поисках их большую территорию.
C. aenescens и Messor denticulatus, труп крупного насекомого, вокруг него моментально собираются муравьи-жнецы. Бегунки обычно держатся поодаль, и время от времени один из них подбегает и пытается оторвать кусок, но моментально отскакивает в сторону, стоит другому муравью прикоснуться к нему антенной или даже резко повернуться. Если говорить об аналогиях с позвоночными, то Cataglyphis скорее можно уподобить классическим некрофагам. Для них характерны приспособления, обеспечивающие возможность контролировать большую территорию, на которой в разных местах случайна появляются источники пищи. На этом аналогии кончаются. Позвоночные обычно питаются трупами крупных животных, которые должны быть расчленены на месте. У них вырабатывается два пути адаптации: постоянное патрулирование территории с малыми затратами энергии (планирование грифов и марабу на большой высоте) или взаимное привлечение на больших расстояниях (характерный вой шакалов и гиен) (Lewic-Goodall, Lewic-Goodall, 1970). Муравьи-бегунки, напротив, обычно собирают трупы, которые могут быть принесены в гнездо одним фуражиром. Каждый фуражир обследует в поисках их большую территорию.
Как и некрофаги-млекопитающие, эти муравьи могут охотиться. Мне неоднократно приходилось наблюдать охоту у Cataglyphis, Proformica и Acantholepis. Cataglyphis и Proformica охотятся всегда в одиночку. Если добыча сравнительно невелика и малоподвижна, муравьи просто берут ее в челюсти и, сжав, несут в гнездо, причем, как правило, схватывают обычно за 1-й или 2-й грудные сегменты. Крупных гусениц эти муравьи кусают, но при сопротивлении сразу же отскакивают в сторону, а затем снова нападают. Это продолжается до тех пор, пока гусеница не перестанет сопротивляться. При охоте на подвижных насекомых (мухи, цикадки) щетинистоногий фаэтончик ( C. setipes) медленно подкрадывается к неподвижной добыче, а затем, когда до нее остается 1—2 см, делает рывок. Если он при этом промахнулся, а жертва перелетела или перепрыгнула на расстояние не более 10—15 см, то он вновь начинает подкрадываться. Обычно подвижные насекомые, на которых охотится фаэтончик, невелики, но в Арчмане (предгорья Копетдага) мне удалось наблюдать охоту этих муравьев на нимф саранчовых, несколько превышающих размер муравья. У Acantholepis semenovi в Копетдаге я также наблюдал охоту на более крупных самцов Tapinoma karavaievi во время брачного лёта последнего вида. Обычно самцы Tapinoma сидят, поводя антеннами и ожидая пробегающих мимо самок, на возвышенных местах (обычно на камнях). Рабочие A. semenovi медленно подкрадываются к ним и затем бросаются и хватают, чаще всего за ноги. Часто при виде начавшейся борьбы подбегают другие рабочие и коллективно убивают жертву. Если поблизости не оказывается других муравьев, жертве всегда удается вырваться. Acantholepis, Proformica и мелкие Cataglyphis питаются в основном трупами. В питании крупных Cataglyphis живые насекомые (крылатые термиты и муравьи-жнецы, гусеницы, мелкие мухи, цикадки) составляют значительную долю, но добывание их, исключая крылатых термитов и муравьев-жнецов во время брачного лёта, несет случайный характер. В то же время сбор трупов муравьев-жнецов фуражирами фаэтончиков — постоянно проводящаяся операция (Длусский, 1975а). Еще более тесная связь наблюдается между степным бегунком ( ![]() C. aenescens) и муравьями-жнецами. Так, в саксаульниках Шафирканского лесхоза ожидаемое среднее расстояние между гнездами бегунка (исходя из плотности гнезд на территории) составляет 98,0 м. Реально среднее расстояние от гнезда этого вида до ближайшего гнезда того же вида — 51,3±9,0 м (n=43), т. е. наблюдается тенденция к агрегации. В то же время среднее расстояние от гнезда M. denticulatus до ближайшего гнезда бегунка — 19,8±б,9 м (n=17). Такая же связь наблюдается между Tetramorium schneideri и мелкими бегунками ( C. emeryi и C. pallida). Подобная ассоциация описана и у американских жнецов Pogonomyrmex и дневного зоонекрофага Conomyrma insana (Wilson, 1971). Остатки пищи C. insana в основном состоят из фрагментов муравьев (Hung, 1974).
C. aenescens) и муравьями-жнецами. Так, в саксаульниках Шафирканского лесхоза ожидаемое среднее расстояние между гнездами бегунка (исходя из плотности гнезд на территории) составляет 98,0 м. Реально среднее расстояние от гнезда этого вида до ближайшего гнезда того же вида — 51,3±9,0 м (n=43), т. е. наблюдается тенденция к агрегации. В то же время среднее расстояние от гнезда M. denticulatus до ближайшего гнезда бегунка — 19,8±б,9 м (n=17). Такая же связь наблюдается между Tetramorium schneideri и мелкими бегунками ( C. emeryi и C. pallida). Подобная ассоциация описана и у американских жнецов Pogonomyrmex и дневного зоонекрофага Conomyrma insana (Wilson, 1971). Остатки пищи C. insana в основном состоят из фрагментов муравьев (Hung, 1974).
Сравнивая поведение пустынных Cataglyphis с поведением родственных им Formica гумидных областей Палеарктики, никак нельзя сказать, что первые, как это считал Вилер, «усилили хищнические привычки». Поведение Cataglyphis больше всего сходно с поведением наименее агрессивных видов из группы ![]() Formica fusca и не идет ни в какое сравнение с поведением настоящих хищников — рыжих лесных муравьев (группа
Formica fusca и не идет ни в какое сравнение с поведением настоящих хищников — рыжих лесных муравьев (группа ![]() Formica rufa).
Formica rufa).
Неоправданным мне кажется и выделение «медовых муравьев» в самостоятельную трофическую группу. Содержимое зоба плерэргатов представляет собой раствор глюкозы, фруктозы и мальтозы с некоторой примесью липидов и глицерина; белков в нем практически не содержится (Burgett, Young, 1974). Поэтому эти муравьи для выкармливания расплода должны добывать ту же пищу, что и другие зоонекрофаги. Действительно, питание степного медового муравья ( Proformica epinotalis), по нашим наблюдениям, не отличается от питания других зоонекрофагов, и в пустынях Казахстана этот муравей замещает Acantholepis semenovi. С другой стороны, тенденция к накоплению меда в зобу определенных рабочих наблюдается у всех пустынных ![]() Formicinae и
Formicinae и ![]() Dolichoderinae. В то же время развиться эта тенденция может лишь при наличии богатых, но не постоянных, источников жидкой сахаристой пищи. В пустынях численность насекомых, выделяющих падь, обычно мала. В. М. Вилер (Wheller, 1908а) указывал, что ночные виды Myrmecocystus, имеющие плерэргатов, распространены в тех местах, где имеется достаточно много деревьев, тогда как в настоящих пустынях встречаются лишь дневные виды, не имеющие этой касты. Правда, большое количество плерэргатов найдено и у дневного пустынного M. melliger (Creighton, Crandall, 1954). В Палеарктике обитают два близких рода, связанных с аридными и семиаридными областями, один из которых ( Proformica) имеет хорошо выраженных плерэргатов, а другой ( Cataglyphis) — не имеет. Сопоставление ареалов этих видов (рис:9) ясно показывает, что Proformica связаны лишь с семиаридными областями и северными пустынями Казахстана и Монголии. Австралийские медовые муравьи Leptomyrmex, вопреки существующему мнению (Wheeler, 1908а, 1915а; Emery, 1920), также не являются обитателями пустынь, что ясно видно при взгляде на ареал этих муравьев (рис:9).
Dolichoderinae. В то же время развиться эта тенденция может лишь при наличии богатых, но не постоянных, источников жидкой сахаристой пищи. В пустынях численность насекомых, выделяющих падь, обычно мала. В. М. Вилер (Wheller, 1908а) указывал, что ночные виды Myrmecocystus, имеющие плерэргатов, распространены в тех местах, где имеется достаточно много деревьев, тогда как в настоящих пустынях встречаются лишь дневные виды, не имеющие этой касты. Правда, большое количество плерэргатов найдено и у дневного пустынного M. melliger (Creighton, Crandall, 1954). В Палеарктике обитают два близких рода, связанных с аридными и семиаридными областями, один из которых ( Proformica) имеет хорошо выраженных плерэргатов, а другой ( Cataglyphis) — не имеет. Сопоставление ареалов этих видов (рис:9) ясно показывает, что Proformica связаны лишь с семиаридными областями и северными пустынями Казахстана и Монголии. Австралийские медовые муравьи Leptomyrmex, вопреки существующему мнению (Wheeler, 1908а, 1915а; Emery, 1920), также не являются обитателями пустынь, что ясно видно при взгляде на ареал этих муравьев (рис:9).
Ни Вилер, ни другие исследователи не выделяют в качестве самостоятельной трофической группы медлительных ночных зоофагов, к которым, в частности, относятся многие пустынные Camponotus. В то же время именно эти медлительные насекомые являются настоящими хищниками и почти не собирают трупов беспозвоночных. Но и эти муравьи не добывают жертв «в угон», а охотятся на малоподвижных насекомых с мягкими покровами. Поразительно упорство этих муравьев во время охоты. Если фаэтончик обычно прекращает преследование после одного—трех неудачных бросков, то Camponotus xerxes может тратить несколько часов на добывание одной жертвы. Хотя ночная активность В пустыне весьма выгодна, переходят к ночной фуражировке представители лишь очень немногих родов муравьев. Скорее всего это связано с трудностями ориентации разведчиков в темноте.
Данные, приведенные в предыдущих разделах, показывают, что во всех пустынях Земли основу коадаптивного комплекса муравьев составляют представители трех трофических групп — дневных зоонекрофагов, ночных зоофагов и карпофагов. Все эти муравьи являются герпетобионтами, т. е. собирают пищу на поверхности почвы. Это вполне естественно, так как для всех пустынь характерна скудная растительность. Поэтому здесь практически отсутствует подстилка, а травяной ярус изрежен и эксплуатируется герпетобионтами. В почвенном ярусе беспозвоночные настолько редки, что обычные для муравьев-геобионтов способы добывания пищи (использование фуражировочных тоннелей) не смогут обеспечить семью питанием. Существенно, что в разных географических районах одни и те же трофические группы часто представлены родами, относящимися к разным трибам и даже подсемействам. При этом они обладают многими, возникшими конвергентно, сходными признаками. Сходство бывает так велико, что еще в начале века в один род объединялись виды, относящиеся в настоящее время к разным трибам, так, например, Myrmecocystus (Lasiini), Cataglyphis (Formicini) и часть Melophorus (Melophorini) или Messor (Pheidolini) и Pogonomyrmex (Myrmicini). Ниже приводятся краткие характеристики этих групп.
1. Дневные зоонекрофаги (рис:10, рис:11).
Особенности биологии. Активны днем в самое жаркое время суток (крупные и средние формы). Питаются трупами насекомых, крылатыми муравьями и термитами, активными днем живыми насекомыми (двукрылые, цикадки, саранчовые, гусеницы и др.). Виды с мелкими и средними размерами рабочих, в основном некрофаги; с увеличением размеров увеличивается доля живой добычи. Большинство видов собирает падь и экссудаты растений и имеет тенденцию к образованию касты плерэргатов. Наиболее ярко эта тенденция проявляется в аридных районах с древесной растительностью и в семиаридных районах.
Черты конвергентного сходства. Виды, рабочие которых имеют мелкие размеры ( Brachymyrmex, Plagiolepis, Iridomyrmex), как правило, относятся к родам, широко представленным в семиаридных и гумидных областях, и не имеют специфических адаптации к жизни в пустынях. У крупных и средних форм обычно развит псаммофор. Грудь (но не голова) довольно сильно вытянута. Конечности длинные. Брюшко компактное; многие виды на бегу задирают его вверх и в этом случае петиолюс имеет форму узелка. Дыхальца узкие и длинные. Иногда тело целиком желтое или покрыто густыми светлыми прилежащими волосками.
Характерные представители:![]() Ponerinae: Ectatommini: Rhytidoponera (Австралия);
Ponerinae: Ectatommini: Rhytidoponera (Австралия);![]() Myrmicinae: Ocymyrmini: Ocymyrmex (Южная Африка);
Myrmicinae: Ocymyrmini: Ocymyrmex (Южная Африка); ![]() Dolichoderinae: Tapinomini: Forelius (Южная Америка), Araucomyrmex (Южная Америка), Dorymyrmex (Южная Америка), Conomyrma (Северная Америка), Tapinoma* (Южная Америка, в пустынях Палеарктики один вид во влажных биотопах, лишь на севере ареала выходящий на плакор), Iridomyrmex* (Австралия, Северная и Южная Америка);
Dolichoderinae: Tapinomini: Forelius (Южная Америка), Araucomyrmex (Южная Америка), Dorymyrmex (Южная Америка), Conomyrma (Северная Америка), Tapinoma* (Южная Америка, в пустынях Палеарктики один вид во влажных биотопах, лишь на севере ареала выходящий на плакор), Iridomyrmex* (Австралия, Северная и Южная Америка);![]() Formicinae: Melophorini: Melophorus (Австралия); Brachymyrmicini: Brachymyrmex* (Южная Америка); Plagiolepidini: Plagiolepis* (Восточная Палеарктика), Anoplolepis (Южная Африка), Acantholepis (Палеарктика); Formicini: Formica (Северная Америка, в Палеарктике в аридных стациях только в Казахстане и Монголии), Proformica (Восточная Палеарктика), Cataglyphis (Палеарктика); Lasiini: Myrmecocystus (Северная Америка, часть видов). /Звездочкой отмечены роды, виды которых, встречающиеся в пустыне, отличаются мелкими размерами (до 3 мм) и не имеют характерных для пустынных зоонекрофагов морфологических адаптации./
Formicinae: Melophorini: Melophorus (Австралия); Brachymyrmicini: Brachymyrmex* (Южная Америка); Plagiolepidini: Plagiolepis* (Восточная Палеарктика), Anoplolepis (Южная Африка), Acantholepis (Палеарктика); Formicini: Formica (Северная Америка, в Палеарктике в аридных стациях только в Казахстане и Монголии), Proformica (Восточная Палеарктика), Cataglyphis (Палеарктика); Lasiini: Myrmecocystus (Северная Америка, часть видов). /Звездочкой отмечены роды, виды которых, встречающиеся в пустыне, отличаются мелкими размерами (до 3 мм) и не имеют характерных для пустынных зоонекрофагов морфологических адаптации./
Особенности биологии. Активны ночью. Охотятся на малоподвижных насекомых с мягкими покровами, появляющихся на поверхности только ночью (термиты), или собирают линяющих насекомых (саранчовых, цикадок). Собирают падь и экссудаты растений. Имеется ярко выраженная тенденция к образованию плерэргатов.
Черты конвергентного сходства. У многих видов развит псаммофор. В остальном часто внешне сходны с видами, обитающими в гумидных областях.
Представители:![]() Formicinae: Camponotini: Camponotus: подрод Tanaemyrmex (Палеарктика, Африка, Австралия и, возможно, Южная Америка), подрод Psammomyrma (Южная Африка), подрод Myrmophyla (Австралия); Lasiini: Melophorus (Северная Америка, часть видов). По-видимому, этот список должен в дальнейшем увеличиться, так как в пустынях Южной Америки, Южной Африки и Австралии никто не занимался изучением биологии муравьев, активных ночью. В частности, весьма возможно, что сюда войдут некоторые Brachyponera
Formicinae: Camponotini: Camponotus: подрод Tanaemyrmex (Палеарктика, Африка, Австралия и, возможно, Южная Америка), подрод Psammomyrma (Южная Африка), подрод Myrmophyla (Австралия); Lasiini: Melophorus (Северная Америка, часть видов). По-видимому, этот список должен в дальнейшем увеличиться, так как в пустынях Южной Америки, Южной Африки и Австралии никто не занимался изучением биологии муравьев, активных ночью. В частности, весьма возможно, что сюда войдут некоторые Brachyponera ![]() (Ponerinae, Ponerini) и Polyrhachis
(Ponerinae, Ponerini) и Polyrhachis ![]() (Formicinae, Camponotini) Австралии.
(Formicinae, Camponotini) Австралии.
В пустынях Северной Америки в качестве ночных зоофагов могут, видимо, выступать и представители рода Novomessor. Однако, поскольку значительную долю в питании этих муравьев составляют семена и поскольку в литературе отсутствуют данные о составе пищи животного происхождения (возможно, они собирают трупы), я включил этот род в число карпофагов.
3. Карпофаги (рис:13, рис:14).
Особенности биологии. Часть видов активны ночью, часть — днем. Основная пища в пустынях — семена растений. Некоторые виды также собирают трупы, живых насекомых и падь, некоторые — облигатные карпофаги. Распространена групповая фуражировка с использованием дорог.
Черты конвергентного сходства. Обычно имеется псаммофор. У многих видов высокая вариабельность рабочей касты, часто наблюдается двухфазная аллометрия, иногда четкий диморфизм. Крупные рабочие с широкими массивными жвалами, с короткими тупыми зубцами. Голова таких рабочих массивная, со вздутыми затылочными углами. У видов, рабочие которых имеют очень мелкие размеры, эти черты обычно не развиты.
Характерные представители:![]() Myrmicinae: Myrmicini: Pogonomyrmex (Северная и Южная Америка); Pheidolini: Messor (Палеарктика, в Южной Африке в семиаридных участках), Oxyopomyrmex (Западная Палеарктика), Veromessor (Северная Америка), Novomessor (Северная Америка), Pheidole (все пустыни), Elasmopheidole (Южная Америка); Solenopsidini: Monomorium (Палеарктика, кроме Центральной Азии, виды рода имеются в пустынях Южной Африки и Австралии, но биология их там не описана), Chelaner (Австралия), Solenopsis (Южная Америка); Tetramoriini: Tetramorium (Палеарктика, представители рода встречаются в пустынях Южной Африки и Австралии, но биология их там не описана).
Myrmicinae: Myrmicini: Pogonomyrmex (Северная и Южная Америка); Pheidolini: Messor (Палеарктика, в Южной Африке в семиаридных участках), Oxyopomyrmex (Западная Палеарктика), Veromessor (Северная Америка), Novomessor (Северная Америка), Pheidole (все пустыни), Elasmopheidole (Южная Америка); Solenopsidini: Monomorium (Палеарктика, кроме Центральной Азии, виды рода имеются в пустынях Южной Африки и Австралии, но биология их там не описана), Chelaner (Австралия), Solenopsis (Южная Америка); Tetramoriini: Tetramorium (Палеарктика, представители рода встречаются в пустынях Южной Африки и Австралии, но биология их там не описана).
Почти всегда в пустынях также встречаются муравьи, которые не могут быть отнесены ни к одной из этих трех основных групп. Во всех случаях это виды родов, широко представленных в сопредельных семиаридных или гумидных областях. Ярко выраженные адаптации к жизни в пустынях у этих муравьев отсутствуют, и часто они сами, либо очень близкие и сходные морфологически виды, могут быть встречены в гумидных условиях. Условно можно выделить следующие четыре группы:
4. Дендробионты-зоофаги.
Особенности биологии. Строят гнезда либо в древесине, либо у корней живых деревьев. Питаются живыми насекомыми в кронах и на стволах, собирают падь и экссудаты растений.
Представители: Pseudomyrmicinae: Pseudomyrmex (Северная и Южная Америка), Tetraponera (Западная Палеарктика, Южная Африка, Австралия);![]() Myrmicinae: Crematogasterini: Crematogaster (Палеарктика, Южная Африка);
Myrmicinae: Crematogasterini: Crematogaster (Палеарктика, Южная Африка);![]() Formicinae: Camponotini: Camponotus (Восточная Палеарктика).
Formicinae: Camponotini: Camponotus (Восточная Палеарктика).
5. Мицетофаги.
Особенности биологии. Питаются грибами, выращенными на субстрате из растительных остатков или экскрементов насекомых.
Представители:![]() Myrmicinae: Attini: Trachymyrmex (Северная Америка), Mycetophylax (Южная Америка).
Myrmicinae: Attini: Trachymyrmex (Северная Америка), Mycetophylax (Южная Америка).
6. Клептобионты.
Особенности биологии. Мелкие муравьи, живущие в гнездах крупных муравьев или термитов. Камеры их гнезд между ходами гнезда хозяина, а галереи соединяются с камерами этого гнезда. По-видимому, поедают трупы, расплод и остатки пищи вида-хозяина. На поверхности появляются только во время брачного лёта.
Представители. Пока известны только один вид Solenopsis в Каракумах, живущий в гнездах термитов, и несколько видов Monomorium из пустынь Южной Африки, живущих в гнездах термитов и муравьев Ocymyrmex. Оба рода относятся к трибе Solenopsidini подсемейства![]() Myrmicinae. Список явно неполон, так как коллектирование (не говоря уже об изучении биологии) клептобионтов представляет большие трудности.
Myrmicinae. Список явно неполон, так как коллектирование (не говоря уже об изучении биологии) клептобионтов представляет большие трудности.
7. Муравьи-кочевники.
Особенности биологии. Не делают постоянных гнезд и постоянно перекочевывают. Численность рабочих в семье от 50 тыс. до 20 млн. Питаются главным образом общественными насекомыми (в пустыне — в основном термитами и муравьями), уничтожая их семьи целиком. По-видимому, эту группу нельзя рассматривать как компонент коадаптивного комплекса муравьев, так как муравьи-кочевники конкурируют не столько с муравьями, сколько с позвоночными-термитофагами и мирмекофагами.
Представители: Dorylinae: Dorylus (Западная Палеарктика, Южная Африка); Ecitinae: Neivamyrmex (Северная Америка).
Каждая из трех основных трофических групп, как правило, во всех пустынях представлена более чем одним видом муравьев. Исключение представляют лишь ландшафты, мало пригодные для их существования, — такыры, пухлые солончаки, бедлендовые пустыни, перевеваемые пески. Возможность одновременного существования на одной территории нескольких видов, относящихся к одной трофической группе, обеспечивается следующими механизмами:
1. Размерная дифференциация. Спектры питания видов, рабочие которых отличаются средними размерами, различны; при разнице в два размерных класса (т. е. по весу в 10 раз) эти спектры практически не перекрываются. Так, средние веса рабочих трех наиболее обычных в песчаной пустыне дневных зоонекрофагов Кызылкумов — Plagiolepis pallescens (ВЗ-1), Cataglyphis pallida (ВЗ-3), C. foreti (ВЗ-5) относятся как 1:8:120. Примерно такое же соотношение весов у некрупной кошки, рыси и тигра. Средний вес самого крупного карпофага Каракумов ( Messor intermedius) в 35 раз больше веса самого мелкого ( Monomorium barbatulum). Хотя этот тип дифференциации достоверно показан нами лишь для пустынь Средней Азии и Дэвидсон и Хансеном для пустынь Северной Америки, можно с уверенностью утверждать, что он очень характерен для всех пустынь Земли. Изучение коллекционных материалов и списков видов локальных фаун показало, что в сборах из одной точки, если они сделаны достаточно полно, всегда имеются крупные, средние и мелкие дневные зоонекрофаги и карпофаги.
2. Поведенческая дифференциация. Пищевые спектры карпофагов одинакового размера могут быть различными, если они используют разную стратегию фуражировки. Муравьи, использующие мобилизацию и фуражирующие с помощью дорог, питаются семенами тех видов растений, которые образуют куртины. Муравьи, фуражиры которых работают как одиночные сборщики, более эффективно собирают семена растений, распределенных дисперсно. Подобные различия в питании были обнаружены Д. Дэвидсон у муравьев-жнецов пустынь Северной Америки и нами у Messor Средней Азии. Некоторые различия в спектрах питания* также сопровождающиеся различиями в способах фуражировки, были обнаружены нами у Cataglyphis foreli и C. setipes в Средней Азии. Первый вид гораздо эффективнее собирает рабочих термитов из разрушенных наземных галерей (лепок), и термиты составляют в его питании большую долю, чем у C. setipes. В большинстве случаев это различие не может обеспечить возможности сосуществования этих видов. Однако в глинистых пустынях Южной Туркмении (Бабадурмаз и др.), где очень высока плотность поселения термитов, эти виды обитают совместно. Можно ожидать, что в тропических пустынях дальнейшее развитие этой тенденции привело к возникновению пищевой специализации и у зоофагов, имеющих одинаковые размеры рабочих, но какие-либо данные на этот счет пока отсутствуют.
3. Пространственная дифференциация. Мелкие муравьи в пределах одного биотопа могут жить и добывать пищу в разных микроассоциациях. Так, в Кызылкумах Tetramorium inerme живет и добывает пищу в кустиках полыни, a Monomorium barbatulum — на открытых местах. Сходные отличия показаны также Витфордом с соавторами для некоторых муравьев Северной Америки. У многих муравьев пустынь Средней Азии наблюдается явная приуроченность к определенному типу почвы. Messor excursionis и Cataglyphis pallida живут на слабо закрепленных песках, M. lamellicornis и C. emeryi — на закрепленных песках и супесях, M. variabilis и C. cinnamomea — на глинистых почвах. Приуроченность к разным типам почв показана также Дели для муравьев Сахары. Данные по другим пустыням отсутствуют, но изучение коллекционных материалов по Melophorus, Chelaner, Ocymyrmex и Dorymyrmex (сравнительные материалы по пустынным Myrmecocystus и Pogonomyrmex в коллекциях СССР отсутствуют) показало, что в пределах этих родов многие виды отличаются по окраске, строению псаммофора и хет передних лапок, что несомненно связано с приспособлением к разным типам почв.
4. Временная дифференциация. В тех пустынях, где травянистые растения вегетируют и плодоносят дважды в год, и средняя температура зимой не ниже 5—10°, разные виды муравьев, обитающие совместно, могут фуражировать в разное время года. Такое явление описано Витфордом с соавторами для Pogonomyrmex и Conomyrma пустыни Чиуауа. Некоторые данные Гринслэйда о динамике активности Melophorus дают основание предполагать, что временная дифференциация имеется и в пустынях Австралии. В пустынях Средней Азии фуражировка всех карпофагов происходит в основном весной. Осенью, правда, созревают семена саксаула и многолетних солянок, но они карпофагами практически не заготавливаются. У зоофагов и зоонекрофагов период фуражировки растянут до осени, но и в этом случае сезонных различий в динамике активности разных видов не наблюдается.
Население муравьев любой пустыни Земли, таким образом, состоит в первую очередь из представителей трех основных трофических групп (дневные зоонекрофаги, ночные зоофаги, карпофаги), причем виды, входящие в одну группу и обитающие на одной территории, как правило, отличаются по размерам рабочих, стациям местообитания или сезонам фуражировки. Почти всегда имеются и какие-либо представители дополнительных групп. Специфический характер местообитания (пустынные деревья, гнезда термитов) или питания (мицетофагия, афидофагия, клептобиоз) позволяет им внедриться без особой конкуренции в уже сложившийся комплекс пустынных видов.
Формально структура коадаптивного комплекса может быть описана как некоторая топологическая структура в многомерном пространстве. Для каждого вида зависимость от каждой из переменных этого пространства описывается вариационной кривой. Соответственно в многомерном пространстве это будет выглядеть как облако, сгущающееся к некоторому центру. Облака всех видов, входящих в комплекс, образуют многомерную топологическую структуру. Необходимым и достаточным будет такое минимальное число переменных, при котором облака ни одной из пар видов не совпадают. Это число будет не особенно большим. Так, для описания комплекса тайги достаточно использовать семь переменных: 1) доля пищи животного происхождения, добываемой каждым видом в кронах деревьев; 2) то же в травяном ярусе; 3) то же на поверхности почвы; 4) то же в подстилке; 5) то же в почве; 6) размер рабочих особей (точнее — размер добычи); 7) плотность особей каждого вида на разных типах почвы. Для описания ядра пустынного комплекса первые пять переменных не являются операциональными, так как ярусное распределение всех видов одинаково. Необходимыми переменными будут: 1) относительная доля семян в питании каждого вида; 2) то же трупов беспозвоночных; 3) то же живых беспозвоночных; 4) относительная доля дисперсно (или компактно) распределенных источников пищи; 5) размер добычи; 6) плотность особей на разных типах почвы; 7) сезонная динамика активности каждого вида.
Минимальное число переменных, необходимое для полного описания структуры комплекса, может служить объективным показателем его сложности. Степень сходства топологических структур может характеризовать степень сходства разных комплексов независимо от того, из каких видов они состоят.
Трудно представить, что столь сходные по своей структуре коадаптивные комплексы муравьев различных пустынь Земли, возникшие на разной основе и представленные неродственными группами муравьев, появились случайно, в результате независимой эволюции каждого из видов. Формирование комплексов — это, несомненно, закономерный процесс, в основе которого лежит длительная коадаптивная эволюция всех или почти всех (возможно внедрение новых видов из семиаридных областей) видов, входящих в комплекс. Пути и закономерности этого процесса будут рассмотрены в заключительной главе после того, как мы рассмотрим, какие адаптации позволяют муравьям существовать в условиях пустынь.
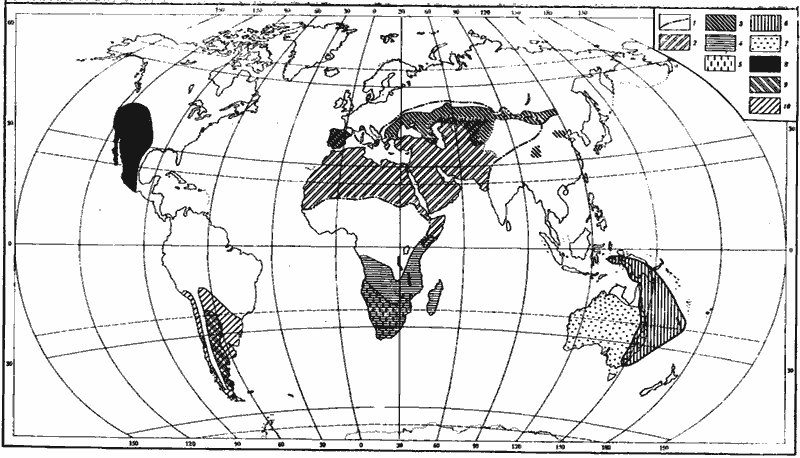
Рис. 9. Географическое распространение основных родов пустынных дневных зоонекрофагов и некоторых «медовых муравьев»
1 — граница распространения Caiaglyphis; 2 — зона, в пределах которой обитает более одного вида Cataglyphis; 3 — Proformica (распространение в КНР, Иране и Турции не изучено; в Афганистане, Индии и Пакистане отсутствуют); 4 — Ocymyrmex; 5 — Anoplolepis (кроме A. longipes, завезенного во многие тропические страны); 6 — Leptomyrmex; 7 — Melophorus; 8 — Myrmecocystus; 9 — Dorymyrmex; 10 — Araucomyrmex

Рис. 10. Пустынные дневные зоонекрофаги (2, 4) в сравнении с близкими видами, обитающими в гумидной зоне (1, 3)
1 —
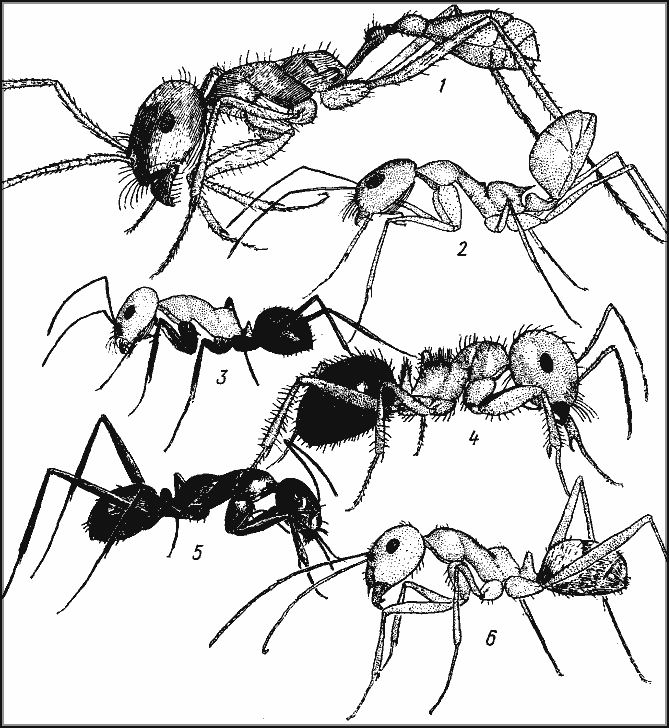
Рис. 11. Дневные зоонекрофаги
1 — Ocymyrmex hirsutus
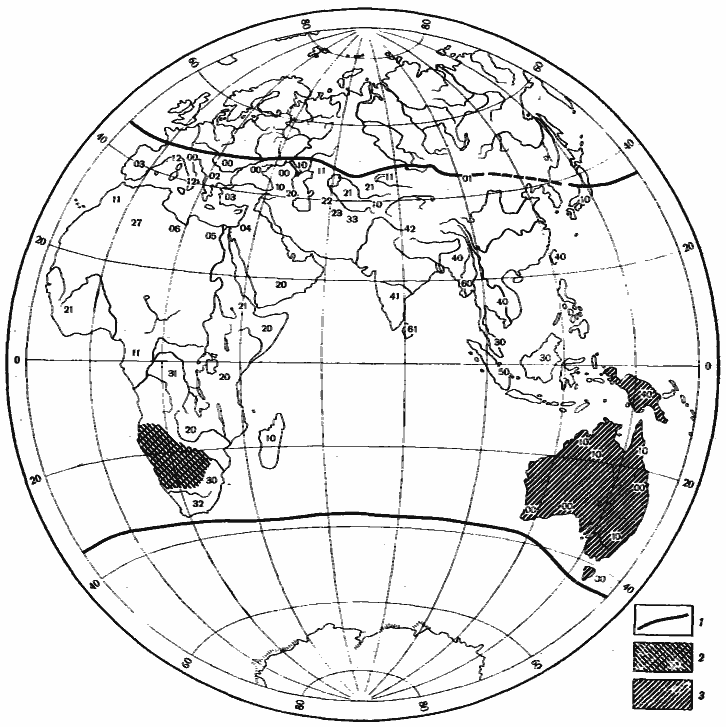
Рис. 12. Некоторые закономерности распространения Camponotus в Восточном полушарии
1 — границы распространения подрода Tenaemyrmex; индекс в пределах ареала подрода: первая цифра — число видов группы C. compressus—sylvaticus (в том числе C. xerxes и C. turcestanus), вторая — группы C. irritans—thraso (в том числе C. turcestanicus и C. fedtschenkoi); 2 — распространение подрода Myrmopsamma; 3 — распространение подрода Myrmophyma (по данным [Emery, 1920] с дополнениями)
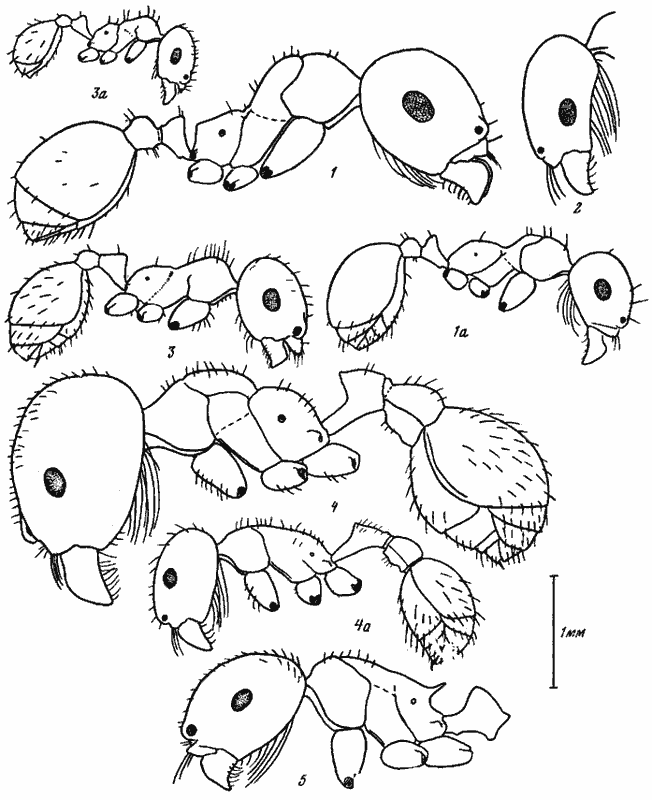
Рис. 13. Муравьи-карпофаги
1, 1a — крупный и мелкий рабочие Messor intermedins (Каракумы); 2 — голова Monomorium chobauti в профиль (Алжир); 3, 3а — Chelaner sp. (Австралия, Дениликуин); 4, 4а — Pogonomyrmex badius (США, Флорида); 5 — P. barbatus molefaciens (США, Техас)
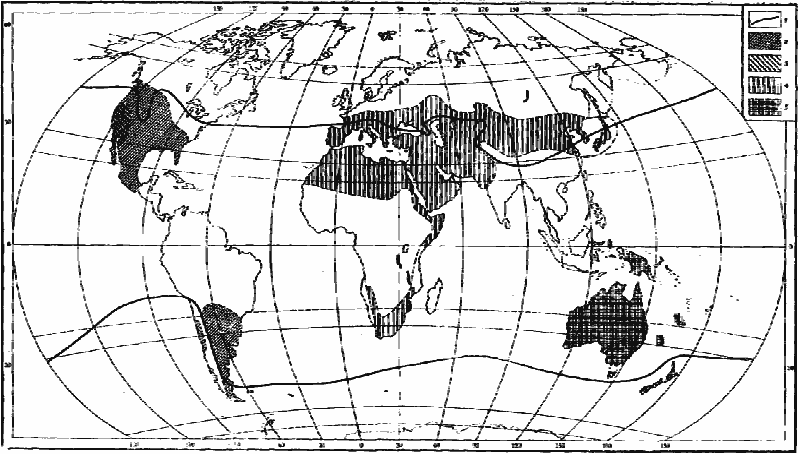
Рис. 14. Распространение некоторых родов
1 — северная и южная границы распространения Pheidole;
2 — Pogonomyrmex;
3 — Veromessor;
4 — Messor;
5 — Chelaner
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
сладкая густая жидкость, выделяемая тлями, червецами и другими насекомыми, питающимися соком растений. Медоносная пчела собирает падь с листьев и перерабатывает в падевый мед.
Всего 30 видов.
Всего 65 видов.
Всего 60 видов.
Всего 106 видов.
Всего 134 вида.
Всего насчитывает 549 видов.
Всего 9 видов. Карпофаги.
Всего 54 вида.
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
у муравьев имеется три основные касты: самцы, самки и рабочие (бесплодные модифицированные самки).
Myrmicinae — самое большое подсемейство муравьев, включающее более 2 тыс. видов, относящихся более чем к 150 родам.
Всего 48 видов.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
 Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунок
Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунокгигрофил, галобионт
Старое название: Cataglyphis flavigastra
гигрофил
(зоонекрофаг), организм, питающийся мертвыми животными, гетеротроф
Всего 24 вида.
ксеробионт
Старые названия: Acantholepis semenovi, Acantholepis transcaspiensis
гигрофил, галобионт
Всего 62 вида.
отряд общественных насекомых. Общины, разделенные на касты, состоят из крылатых и бескрылых особей. Строят подземные и наземные (до 15 м высотой) гнезда (термитники). Ок. 2600 видов, главным образом в тропиках; в России 2 вида: один в районе Сочи, второй, вероятно, завезенный из Японии или Китая, во Владивостоке. Многоядны. Разрушают древесину, портят бумагу, кожу, сельскохозяйственные продукты и др. Термиты, обитающие в почве, полезны как почвообразователи.
ксеробионт
Старое название: Proformica deserta
Старое название: Conomyrma insana
 Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant
Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравейрабочие с сильно раздутым брюшком; это каста, так называемых, "медовых бочек".
Одно из крупных подсемейств, насчитывает около 1,5 тыс. видов, относящихся к 44 родам. Широко распространены на всем земном шаре, довольно обычны в тропиках и занимают доминирующее положение в умеренных широтах.
Небольшое наиболее архаичное подсемейство муравьев. Распространено по всему земному шару, преимущественно в тропиках. Насчитывает более 230 видов.
Всего 17 видов.
организм, питающийся исключительно другими животными
Всего 965 видов.
животные, не имеющие позвоночника: простейшие, губки, кишечнополостные, черви, моллюски, членистоногие, иглокожие и др.; примерно 3/4 видов всех животных.
организм, питающийся семенами растений.
организм, обитающий среди растительных или иных органических остатков, на поверхности почвы.
организм, обитающий на поверхности и в толще пород суши.
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
Всего 23 вида.
Всего 39 видов.
Всего 60 видов.
Всего 58 видов.
специальное приспособление для переноски сухой почвы, распространенн у пустынных муравьев, образован длинными отстоящими волосками (аммохетами), часто имеющими форму латинской буквы J, расположен на нижней поверхности головы.
особый сегмент брюшка. У муравьев бывает двучлениковый (петиолюс + постпетиолюс) и одночлениковый (петиолюс - чешуйка). Количество члеников и форма стебелька очень важны для идентификации вида. Например у муравьев семейства Formicinae стебелек одночлениковый, а у Myrmicinae - двучлениковый.
Довольно примитивная группа муравьев по своему строению и поведению. Рабочие мономорфны, самки по размерам близки к рабочим.
Всего 114 видов.
Всего 17 видов.
(гр. topos - место) - естественное жизненное пространство определенного биоценоза, совместно с которым он образует экосистему.
«медовые муравьи»
Всего 22 вида.
Всего 477 видов.
Всего 172 вида.
Включает сокращенный Novomessor.
итал. obligato, "обязательный", необходимый, существенный
(лат. mandibule — челюсть, от mando — жую, грызу), первая пара челюстей у ракообразных, многоножек и насекомых. Синоним: жвала.
Всего 415 видов
организм, обитающий в древесном ярусе растительности.
Всего 129 видов.
Всего насчитывает 85 видов.
Всего 427 видов.
организм, питающийся мицелием грибков
Всего 42 вида.
организм, питающийся за счет воровства
вид, семьи которого используются муравьями - социальными паразитами для основания семьи или постоянного обитания.
живут колониями (пчелы, муравьи, термиты, некоторые осы). В состав колонии, напр. у термитов, входят половые особи (самцы и матки обеспечивают размножение), солдаты (защиту) и рабочие (собирание корма, строительство гнезда и т. д.). Солдаты и рабочие бесплодны; половые особи и солдаты сами не способны питаться; таким образом, члены колонии полностью зависят друг от друга. Разделение функций и сложные взаимодействия разных особей в колониях общественных насекомых обеспечиваются рядом особенностей в их строении и поведении; последнее регулируется с помощью выделяемых ими химических веществ (феромонов).
организм, питающийся муравьями
Всего 61 вид.
Всего 126 видов.
олигогинны
индивидуальное или групповое привлечение пассивных фуражиров активными фуражирами для конкретных полей в конкретном месте.
ксеробионт
Старое название: Cataglyphis cinnamomea
организм, питающийся тлями (Aphididae) и их паразитоидами
необратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.
Старое название: Anoplolepis longipes
 Rhytidoponera metallica (Smith, 1858) — зелёноголовый муравей
Rhytidoponera metallica (Smith, 1858) — зелёноголовый муравейорганизм, обитающий в лесной подстилке.
Старое название: Araucomyrmex tener






