






Общее расчленение тела. Тело муравьев делится на голову, мезосому или грудь, стебелек и брюшко. Грудь (мезосома) муравьев к других Apocryta не гомологична груди других насекомых, поскольку образована путем слияния торакса и первого абдоменального сегмента. Стебелек, образующий с грудью шарнирное, а с брюшком — шарнирное, или кольцевое подвижные сочленения, может состоять из одного или двух члеников. Первый, часто единственный, членик стебелька называется петиолюсом, второй — постпетиолюсом. Они гомологичны, соответственно, II и III истинным абдоменальным сегментам. У видов с одночлениковым стебельком брюшко самок и рабочих состоит из 5, а самцов — из 6 сегментов, гомологичных, соответственно, III—VII и III—VIII истинным абдоменальным сегментам. У муравьев с двучленнковым стебельком число сегментов на один меньше и, соответственно, I брюшной сегмент гомологичен IV истинному. Стебелек и брюшко вместе образуют метасому. Сегментация тела у представителей разных подсемейств показана на рис:6.
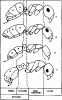
Рис. 6. Расчленение тела рабочих муравьев (ориг.):
А -
Б - Tapinoma erraticum (
В - Ponera coarctata
Г - Myrmica bessarabica
Голова. Голова муравьев (рис:9) состоит из головной капсулы, несущей пару усиков (антенн), фасеточные глаза, простые глазки и ротовые части. Фасеточные глаза обычно имеются у всех трех каст, но у рабочих особей некоторых видов (Ponera, Diplorhoptrum и др.) они могут быть редуцированы до нескольких фасеток, а у Leptanillinae и большинства Dorylinae как рабочие, так и самки полностью лишены глаз и простых глазков. Последние всегда имеются у крылатых особей, а у рабочих их наличие или отсутствие является важным диагностическим признаком.
На головной капсуле почти всегда хорошо виден клипеус (наличник), отделенный швом. От верхнего края его отходят лобные валики; часто в передней части они развиваются в лобные лопасти, частично или полностью прикрывающие места прикрепления усиков, находящиеся в усиковой ямке. У некоторых муравьев (например у Camponotus) на шве клипеуса имеется также обособленная клипеальная ямка в том месте, где к капсуле изнутри прикреплена ветвь фурки внутреннего скелета головы, но в большинстве случаев клипеальная и усиковая ямки слиты. Между лобными валиками к заднему краю клипеуса примыкает треугольная лобная площадка, обычно также ограниченная швами. Иногда между лобной площадкой и клипеусом шов исчезает, и они сливаются.
Рис. 7. Мускулатура головы Aphaenogaster raphidiiceps (а) и Messor aralocaspius (б) (Длусский, 1981): f - головная фурка; g - гулярный шов; mem - мышцы открывающие мандибулы; mfm - мышцы, сжимающие мандибулы.
Различают также следующие условные поверхности: лоб — между лобными валиками, задним краем клипеуса и областью простых глазков; затылок — между затылочными углами, областью глазков и затылочным отверстием; щеки — боковая поверхность головы между основанием мандибул и нижним краем глаза; виски — боковая поверхность головы между верхним краем глаза и затылочными углами; нижняя поверхность головы.
Усики самок и рабочих 4—12-члениковые, всегда коленчатые. Первый членик носит название скапус, остальные вместе взятые — жгутик. Длина скапуса составляет 30—45% от длины всего усика. Последние 2—4 членика иногда бывают утолщенными и образуют булаву (рис:8). Усики самцов 12—13-члениковые, причем 12-члениковые — только у видов, самки которых имеют 11 и менее члеников усика. Усики самцов могут быть коленчатыми или нитевидными, с коротким скапусом, длина которого может быть даже меньше длины 2 или 3 члеников. Форма 2-го членика (педицеллюса) у![]() Myrmicinae — важный родовой признак.
Myrmicinae — важный родовой признак.
Ротовой аппарат лижуще-грызущего типа состоит из непарной верхней губы (лябрум), парных мандибул и максилл и непарной нижней губы («язычка»), образовавшейся из слияния парных элементов. Мандибулы у большинства видов треугольные с развитым зазубренным жевательным краем. У некоторых видов (Epitritus, Amblyopone) они длинные с редкими зубцами, а у некоторых муравьев-рабовладельцев (Polyergus, Strongylognathus) могут иметь вид клыков и лишены зубцов. У самцов некоторых видов мандибулы сильно редуцированы. Строение максилл и нижней губы неодинаково в разных группах муравьев, но в систематике эти признаки практически не используются, поскольку детали строения видны только на специально приготовленных препаратах. Лишь в некоторых случаях используются такие признаки, как число и соотношение длины члеников максиллярных и нижнегубных щупиков.
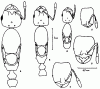
Рис. 8. Тело самок сверху (а, в, д) и голова рабочих сверху (б, г, е) Diplorhoptrum(ориг.):
а, б -
в, Г - D. deserticolum;
д, е - D. celatum (д - голотип).
Форма головной капсулы функционально связана со строением мандибул, поскольку мышцы, сдвигающие их крепятся своими основаниями в затылочных углах головы. У видов со слабо развитыми мандибулами, снабженными острыми зубцами, т.е. неспециализированных хищников (например Aphaenogaster; см. рис:7/а/), эти мышцы развиты слабо, и голова имеет яйцевидную форму. У видов, питающихся твердыми семенами (Messor, солдаты Pheidole (см. рис:7/б/)) или строящих гнезда в древесине (Liometopum, солдаты Camponotus, ![]() Lasius fuliginosus) мандибулы массивные, часто с затупленными зубцами, объем мышц увеличен, а затылочные углы вздуты и часто затылочный край вогнут. Большинство наших муравьев имеет промежуточное между этими крайними типами строение и голова у них имеет более или менее прямоугольную форму.
Lasius fuliginosus) мандибулы массивные, часто с затупленными зубцами, объем мышц увеличен, а затылочные углы вздуты и часто затылочный край вогнут. Большинство наших муравьев имеет промежуточное между этими крайними типами строение и голова у них имеет более или менее прямоугольную форму.
Грудь (мезосома). Грудь муравьев состоит из переднегруди, среднегруди и проподеума (в старой литературе часто использовался термин эпинотум), образовавшегося из слившихся заднегруди и I брюшного сегмента (рис:9). Грудь самок и самцов расчленена сравнительно полно, во всяком случае столь же полно, как у других Apocryta, и несет 3 пары ног и 2 пары крыльев, которые самки сбрасывают после оплодотворения. Рабочие особи не имеют крыльев и их грудь сильно видоизменяется. В пределах семейства прослеживается явная тенденция к слиянию склеритов и исчезновению шва.
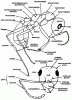
Рис. 9. Строение тела муравьев (ориг.):
вверху - рабочий
внизу - голова рабочего Formica subpilosa (сверху).
Переднегрудь представлена только пронотумом (переднеспинкой). Плейриты переднегруди слились с шейным отделом, к которому неподвижно прикреплен рудимент стернита. Шейный отдел подвижно сочленен с пронотумом. У некоторых муравьев последний также обладает небольшой подвижностью относительно среднегруди. У самок и самцов среднегрудь расчленена более полно, чем остальные отделы, и состоит из мезонотума (среднеспинки), расчлененного, в свою очередь, на скутум и скутеллюм, мезоплевр, в свою очередь расчлененных на эпимерит и эпистернит, и метастернита. От передних углов скутума у некоторых муравьев отходят отчетливые парапсидальные швы (бороздки Майра). На нижней поверхности среднегруди, частично сохраняются швы, отделяющие мезостернит от мезоплевритов. У самок и самцов также сохраняется отделенный швами метанотум, за которым следует проподеум, образованный слиянием метаплевритов, метастернита и I брюшного сегмента. Профиль проподеума часто бывает более или менее угловатым. В этом случае различают его основную (верхнюю) и покатую (от угла до места прикрепления петиолюса) поверхности. Мезосома рабочих особой устроена проще. Даже в случае наиболее полного расчленения сохраняются лишь швы, отделяющие пронотум от мезонотума (промезонотальный), мезонотум от мезоплевр (мезоплевральный), и среднегрудь от проподеума (мезопроподеальный), причем у некоторых муравьев (Tapinoma, Messor) проподеум несколько подвижен относительно среднегруди. В пределах семейства наблюдается явная тенденция к дальнейшему слиянию грудных швов у рабочих особей, и у некоторых муравьев швы полностью исчезают. Лишь шейный отдел во всех случаях отделен от остальной части и груди и относительно подвижен.
На мезопроподеальном шве и на проподеуме имеются дыхальца. Форма проподеального дыхальца в ряде случаев является важным диагностическим признаком. В нижних задних углах проподеума открываются отверстия метаплевральных желез; обычно также хорошо заметно вздутие над резервуаром этих желез. Часто проподеум несет парные зубцы или шипы, форма которых у![]() Myrmicinae является важным таксономическим признаком.
Myrmicinae является важным таксономическим признаком.
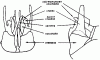
Ноги состоят из коксы, вертлуга, бедра, голени и 5-члениковой лапки с двумя коготками на последнем членике. Между коготками у некоторых муравьев имеется аролий. Передняя нога отличается от остальных тем, что на ней имеется специальное приспособление для чистки усиков и лапок, образованное изогнутыми вершиной голени и основанием 1-го членика лапки и большой изогнутой гребенчатой шпорой (чистильный аппарат). На вершинах средней и задней голеней у примитивных муравьев имеется по две пары шпор, одна из которых простая, а другая гребенчатая. У большинства наших муравьев одна или обе эти шпоры редуцированы.
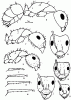
Рис. 11. Тело в профиль (А,Г,З), голова сверху (Б,Д,Е,И) и хетотаксия задней голени (В,Ж,К,Л) солдат и рабочего (Е) представителей подрода Tanaemyrmex рода Camponotus (ориг.): А,Б,В - C. turkestanus - Г-Ж - C. fedtschenkoi; 3,И,К - C. obliquipilosus; Л - C. bactrianus.
В систематике муравьев используется только жилкование переднего крыла, причем наличие или отсутствие тех или иных ячеек может быть важным родовым признаком, но на видовом уровне признаки жилкования не используются. Наиболее полное жилкование переднего крыла имеется у большинства![]() Ponerinae (наши Ponera и Amblyopone) и некоторых
Ponerinae (наши Ponera и Amblyopone) и некоторых ![]() Dolichoderinae. У этих муравьев имеются замкнутые радиальная, 1-я и 2-я радиомедиальные (кубитальные) и дискоидальная (медиокубитальная) ячейки. У большинства наших муравьев наблюдается большая или меньшая редукция этого исходного типа (рис:49).
Dolichoderinae. У этих муравьев имеются замкнутые радиальная, 1-я и 2-я радиомедиальные (кубитальные) и дискоидальная (медиокубитальная) ячейки. У большинства наших муравьев наблюдается большая или меньшая редукция этого исходного типа (рис:49).
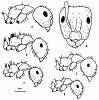
Рис. 12. Солдаты представителей подрода Myrmentoma рода Camponotus (ориг.);
А,Б - C. lameerei;
В - C. semirufus;
Г - C. interjectus;
Д - C. kopetdaghensis (паратип);
Е - C. lateralis.
Метасома (см. рис:9). У муравьев с одночлениковым стебельком тергит петиолюса развивается в поперечную лопасть, носящую название чешуйки. Чешуйка может быть вертикальной (см. рис:3, рис:11, рис:12, рис:13, рис:14, рис:15) или наклоненной вперед (см. рис:4 и рис:16). Реже петиолюс может быть цилиндрическим (Tapinoma) или узловидным (некоторые Cataglyphis, (рис:17/е,д/)). Напротив, у видов с 2-члениковым стебельком форма петиолюса бывает, чаще всего, узловидной, с развитым узелком различной формы и, как правило, удлиненной передней цилиндрической частью (см.рис:6/Г/, рис:8, рис:18—33), реже цилиндрической (Myrmecina). Постпетиолюс у этих видов имеет более или менее шаровидную форму. Своеобразная форма петиолюса у примитивных Amblyoponini: он отделен от брюшка сравнительно слабой перетяжкой и, если смотреть сверху, прикреплен к нему всей задней поверхностью.
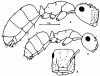
Первый тергит брюшка![]() Ponerinae,
Ponerinae,![]() Formicinae и
Formicinae и ![]() Dolichoderinae, гомологичный постпетиолюсу
Dolichoderinae, гомологичный постпетиолюсу![]() Myrmicinae, имеет форму куба без двух сторон, с сильно округленными углами и гранями. Его верхняя поверхность называется основной, а передняя — покатой. У
Myrmicinae, имеет форму куба без двух сторон, с сильно округленными углами и гранями. Его верхняя поверхность называется основной, а передняя — покатой. У![]() Ponerinae I сегмент брюшка отделен от II перетяжкой. Эти сегменты имеют шаровидное сочленение, образованное жестким хитиновым пояском на переднем крае II сегмента и суженно заднем краем I. В остальных случаях сочленения между брюшными сегментами трубчатые, т.е. последние способны не только вращаться относительно друг друга, но и втягиваться друг в друга.
Ponerinae I сегмент брюшка отделен от II перетяжкой. Эти сегменты имеют шаровидное сочленение, образованное жестким хитиновым пояском на переднем крае II сегмента и суженно заднем краем I. В остальных случаях сочленения между брюшными сегментами трубчатые, т.е. последние способны не только вращаться относительно друг друга, но и втягиваться друг в друга.
Рис. 14. Рабочий Polyrhachis simplex (Афганистан). Слева - тело в профиль, справа - грудь и петиолюс сверху (ориг.).
У самок и рабочих особей![]() Ponerinae и
Ponerinae и![]() Myrmicinae имеется жало, гомологичное яйцекладу. Строение его в разных группах муравьев может быть различным (Kugler, 1918), но в систематике эти признаки практически не используются. У
Myrmicinae имеется жало, гомологичное яйцекладу. Строение его в разных группах муравьев может быть различным (Kugler, 1918), но в систематике эти признаки практически не используются. У![]() Formicinae,
Formicinae, ![]() Dolichoderinae жало редуцировано, хотя железы ядовитого аппарата развиты порой сильнее, чем у видов, имеющих жало. Секрет желез эти муравьи выбрызгивают, причем иногда на расстояние в несколько десятков сантиметров. У
Dolichoderinae жало редуцировано, хотя железы ядовитого аппарата развиты порой сильнее, чем у видов, имеющих жало. Секрет желез эти муравьи выбрызгивают, причем иногда на расстояние в несколько десятков сантиметров. У ![]() Dolichoderinae отверстие желез скрыто под стернитами брюшка. У
Dolichoderinae отверстие желез скрыто под стернитами брюшка. У![]() Formicinae выход ядовитой железы вытянут в трубочку (ацидопору), окруженную венчиком волосков (коронулой) (см. рис:6/А/).
Formicinae выход ядовитой железы вытянут в трубочку (ацидопору), окруженную венчиком волосков (коронулой) (см. рис:6/А/).
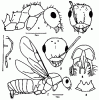
Рис. 15. Alloformica (ориг.): А,Б - A. obscurior sр.nov. (голотип) (А - тело в профиль, Б - голова сверху); В - максиллярный щупик рабочего A. aberrans; Г-Ж - самец A. aberrans (андртип) (Г - тело в профиль, Д - голова сверху, Е - гениталии сверху, Ж - субгенитальная пластинка снизу).
Гениталии самцов (см. рис:10) сострят из субгенитальной пластинки и трех пар створок. Наружная створка состоит из двух частей — сквамулы и стипеса. Часто на них имеются различной формы выросты. Внутренние створки (сагитты) широкие и обычно зазубренные с брюшной стороны. Между стипесами и сагиттами помещаются вольселлы сложной формы, часто с крючковидным выростом на конце. В некоторых родах, например, Tapinoma, Cataglyphis, строение гениталий самцов — важный диагностический признак, в других — например у Formica, их строение у разных видов почти не отличается.
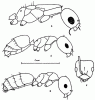
Рис. 16. Представители рода Plagiolepis (ориг.): а,б - Pl. pallescens (а - самка, б - рабочий); в,г - Pl. arnoldii sp.n. (голотип) (в - рабочий в профиль; г - голова того же экземпляра сверху).
Окраска, скульптура, опушение. Окраска муравьев не так бросается в глаза, как у многих других жалящих перепончатокрылых. Правда, в тропиках некоторые виды имеют металлическую окраску, но наши муравьи черные, бурые, коричневые, желтые, буровато- или желтовато-красные. Часто грудь и голова бывают буровато- или желтовато-красными, брюшко — бурым или черным.

Рис. 17. Крупные виды Cataglyphis (ориг.): а,б - грудь в профиль и максиллярный щупик C. foreli (топотип); в - C. murgabica (паратип; Имам-Баба); г,д - крупный и мелкий рабочие
Важное значение в систематике муравьев, особенно на видовом уровне, имеет скульптура различных частей тела. Она может и отсутствовать, тогда поверхность бывает гладкой и блестящей. Грубая скульптура видна даже при небольших увеличениях, а поверхностная заметна только при большом увеличении и направленном косом освещении. Естественно, четких границ между этими двумя типами не существует. Грубая скульптура, состоящая из изолированных углубленных ямок, называется ямчатой, а поверхностная — точечной. Грубая скульптура,состоящая из более или менее параллельных или расходящихся морщинок, называется морщинистой, а поверхностная — струйчатой. Грубая скульптура из морщинок, образующих сеть замкнутых ячеек неправильной формы, называется ячеистой, а поверхностная — сетчатой. Грубая скульптура из соприкасающихся бугорков называется гранулированной, а поверхностная — шагреневой.
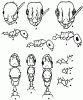
Рис. 18. Представители рода Temnothorax
(Длусский, Союнов, 1988). Голова рабочих сверху (а,д,и), тело рабочих в профиль (б,е,к), тело самок сверху (в,ж,л), стебелек и проподеум самок в профиль (г,з,м); а-г - T. recedens, д-з - T. semenovi, и-м - T. desertorum (голотип и паратип).
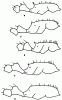
Рис. 19. Грудь и стебелек рабочих Leptothorax (ориг.):
а - L. unifasciatus (Копетдаг); б - L. melleus (паратип, Шугнан); в - L. parvulus (Кара-Кала); г - L. hasardaghi (паратип, Ел-Дере); д - L. anodontoides (паратип, Ай-Дере).
Важным таксономическим признаком для систематики муравьев также является строение, расположение и обилие волосков. Различают отстоящие и прилегающие волоски. Отстоящие волоски (хеты) обычно крупные, жесткие и в основании их имеется волосковая ямка. Прилегающие волоски обычно плотно прилегают к телу и образуют прилежащее опушение. Хотя в большинстве случаев эти группы волосков хорошо отличаются, могут быть и промежуточные состояния. Прилежащие волоски могут быть приподнятыми, наклонными и полуотстоящими (т.е. наклоненные под углом 45°). При описании прилежащего опущения соотносят длину волосков с расстоянием между ними, а при описании хетотаксии указывают приблизительное число волосков, которые видны при рассматривании в стандартном положении. Для описания хетотаксии груди, стебелька и брюшка тело рассматривается точно в профиль (см. рис:9, рис:11). Число волосков на затылочном крае головы, висках и щеках подсчитывается при рассмотрении головы точно сверху (см. рис:9, рис:11 и др.).
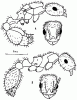
Особо следует оказать о псаммофорах — специальных приспособлений для переноски сухой почвы, распространенных у пустынных муравьев. У представителей подсемейства![]() Myrmicinae и у Camponotus псаммофор образован длинными отстоящими волосками (аммохетами), часто имеющими форму латинской буквы J, расположенной на нижней поверхности головы (рис:34/Б/). При несовершенном псаммофоре эти волоски обычно почти прямые и расположены беспорядочно. Совершенный псаммофор образован J-образными волосками, расположенными правильными рядами. Специализированный псаммофор представляет собой площадку, лишенную волосков, окруженную одним рядом длинных J-образных волосков, образующих корзинку (см. рис:30/д/). Особое строение имеет псаммофор Cataglyphis. Его задняя стенка образована длинными волосками на нижней стороне головы, дно — длинными нижнечелюстными щупиками, бока — мандибулами, несущими длинные волоски, а передняя стенка — длинными жесткими хетами на переднем крае клипеуса (см. рис:34/А/).
Myrmicinae и у Camponotus псаммофор образован длинными отстоящими волосками (аммохетами), часто имеющими форму латинской буквы J, расположенной на нижней поверхности головы (рис:34/Б/). При несовершенном псаммофоре эти волоски обычно почти прямые и расположены беспорядочно. Совершенный псаммофор образован J-образными волосками, расположенными правильными рядами. Специализированный псаммофор представляет собой площадку, лишенную волосков, окруженную одним рядом длинных J-образных волосков, образующих корзинку (см. рис:30/д/). Особое строение имеет псаммофор Cataglyphis. Его задняя стенка образована длинными волосками на нижней стороне головы, дно — длинными нижнечелюстными щупиками, бока — мандибулами, несущими длинные волоски, а передняя стенка — длинными жесткими хетами на переднем крае клипеуса (см. рис:34/А/).
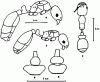
Рис. 21. Самки (а,г), петиолюс самок сверху (в,д) и рабочий (б) Cardiocondyla koshewnikovi (а-в) и C. elegans (г,д) (ориг.).
Органы чувств. Органы зрения муравьев представлены фасеточными глазами и тремя простыми глазками, функция последних пока не очень ясна, но несомненно, что у муравьев они имеют какое-то отношение к полету. Они имеются и хорошо развиты у крылатых особей всех видов, тогда как у бескрылых самок Dorylinae и Leptanillinae и рабочих либо отсутствуют, либо сильно редуцированы. Степень различий в развитии фасеточных глаз у бескрылых особей (у крылатых они всегда хорошо развиты) в разных группах муравьев очень велика. У южно-американских Gigantiops огромные глаза занимают почти всю боковую поверхность головы. Достаточно сильно развиты глаза и у наших Formica или Cataglyphis. с другой стороны, у Ponera или Diplorhoptrum они редуцированы до одной или нескольких фасеток, а у Dorylinae, Leptanillinae и некоторых![]() Myrmicinae полностью отсутствуют.
Myrmicinae полностью отсутствуют.
Рис. 22. Паразитические
Зрение муравьев электрофизиологическими методами изучали Мазохин-Поршняков и Тренн (1972) на Cataglyphis, Formica и Lasius, а поведенческими — Мурзин (1976) на ![]() Formica sanguinea. Показано, что разрешающая способность глаза у исследованных муравьев ниже, чем у человека или медоносной пчелы, но муравьи различают отдельные вспышки, подаваемые с частотой до 70 гц. Муравьи обладают цветовым зрением, но оно у них бихроматичеокое, а не трихроматическое, как у человека или медоносной пчелы. Как и медоносная пчела, муравьи способны воспринимать направления колебаний поляризованного света.
Formica sanguinea. Показано, что разрешающая способность глаза у исследованных муравьев ниже, чем у человека или медоносной пчелы, но муравьи различают отдельные вспышки, подаваемые с частотой до 70 гц. Муравьи обладают цветовым зрением, но оно у них бихроматичеокое, а не трихроматическое, как у человека или медоносной пчелы. Как и медоносная пчела, муравьи способны воспринимать направления колебаний поляризованного света.
Хотя у многих![]() Myrmicinae, в том числе наших Myrmica, имеется стридуляционный аппарат, образованный задним краем постпетиолюса и. поперечными морщинками на переднем крае первого брюшного тергита (Markl et al., 1968; Жантиев, Сулханов, 1977), специализированные органы слуха у них, видимо, отсутствуют. Как показали Mark et al. (1968), муравьи воспринимают не звук, а колебания почвы, но это не мешает им реагировать на стридуляцию других муравьев.
Myrmicinae, в том числе наших Myrmica, имеется стридуляционный аппарат, образованный задним краем постпетиолюса и. поперечными морщинками на переднем крае первого брюшного тергита (Markl et al., 1968; Жантиев, Сулханов, 1977), специализированные органы слуха у них, видимо, отсутствуют. Как показали Mark et al. (1968), муравьи воспринимают не звук, а колебания почвы, но это не мешает им реагировать на стридуляцию других муравьев.
Органами осязания служат отстоящие волоски (сенсиллы), расположенные на последних члениках усика. При помощи отстоящих волосков муравьи также воспринимают движения воздуха и сотрясение субстрата.

Рис. 24. Aphaenogaster messoroides sp. nov. (ориг.): а - самец (паратип); б - голова самца; в - рабочий (голотип); г - голова голотипа.
В различных местах на теле муравьев также имеются участки, густо покрытые полосками (поля щетинок). Как показал Hubert (1962), они являются рецепторами силы тяжести. При горизонтальном движении ориентация осуществляется за счет коксальных и брюшных полей щетинок, а при вертикальном — шейных, петиолярных, антеннальных и коксальных. Путем последовательного исключения полей показано, что для правильной ориентации должна быть подвижной хотя бы одна из систем рецепторов.

Рис. 25. Рабочие Messor из групп barbatulum (а–б) и aralocaspius (ж–к) (ориг.): а - M. intermedius, тело в профиль; б - он же, голова сверху; в - M. denticulatus, тело в профиль; г - он же, голова сверху; д - он же, основание скапуса; е - он же, 3 первые членика жгутика; ж - M. aralocaspius тело в профиль; з - он же, голова сверху; и - M. subgracilinodis (паратип), тело в профиль; к - он же, голова сверху.
Обоняние развито у муравьев очень хорошо. Многочисленные обонятельные сенсиллы располагаются на последних члениках жгутика усика. У некоторых муравьев эти членики увеличены и образуют булаву. Часто это сопровождается редукцией глаз, что указывает на то, что муравьи с развитой булавой (как правило, это виды, охотящиеся в почве и подстилке) больше руководствуются обонянием и осязанием, чем зрением. Forel (1921) обратил внимание на то, что органы обоняния разнесены у муравьев (как впрочем и у других насекомых)на довольно большое расстояние, так что запах у этих насекомых должен быть объемным, подобно зрению и слуху у позвоночных животных.
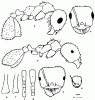
Рис. 26. Рабочие Messor из группы structor (ориг.); а–г –
Половая система. Половой аппарат самцов состоит из парных семенников, парных семенных протоков, которые сливаются в непарный проток, открывающийся в эдеагус. Половой аппарат самок состоит из большого числа яйцевых трубочек, открывающихся в парные яйцеводы, которые, сливаясь, образуют непарный яйцевод. Семеприемник служит для хранения спермы, которую самки муравьев сохраняют в течение всей жизни, так как оплодотворение у них происходит лишь один раз в жизни. Семеприемник снабжен специальной парной железой и открывается в непарный яйцевод. У рабочих особей многих видов муравьев, как и у самок, имеются яйцевые трубочки, но если у самок их число колеблется от нескольких десятков до нескольких сотен, то у рабочих — обычно не превышает 10. Во многих случаях яичники видны только у молодых рабочих.

Рис. 27. Рабочие Messor из группы vicinus (ориг.): а-г - M. perantennatus; д-з - M. vicinus (а, д - тело в профиль; б,е - 3 первые членика жгутика; в,ж - основание скапуса; г,з - голова сверху).
Железы. Выделения желез внешней секреции имеют в жизни муравьев важное значение. Помимо довольно простых первичных функций, эти железы часто приобретают вторичные, связанные с общественным образом жизни. Выше уже говорилось, что секреты лабиальных и фарингеальных желез, помимо первичных функций (выделение ферментов), у некоторых муравьев служат для выкармливания личинок. Возможно, они также используются и при постройке картонных гнезд. Некоторые тропические муравьи используют выделения паутинных желез личинок для постройки гнезд.

Рис. 28. Рабочие Messor из группы excursionis (ориг.): а-в - M. excursionis; г-е - M. lamellicornis; ж-и - M. variabilis. (а,г,ж - тело в профиль; б,д,з - голова сверху; в,е,и -основание скапуса).
У общественных насекомых выделения различных желез служат феромонами, т.е. сигнальными веществами, влияющими на поведение других особей своего вида. Феромоны и феромонные железы муравьев исследованы достаточно хорошо (Wilson, 1971; Holldobler, Engel, 1978; Брайен, 1983), но наиболее изучены следовые феромоны, т.е., вещества, при помощи которых муравьи метят тропы, и феромоны тревоги, вызывающие агрессивную реакцию семьи.
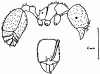
Первичная функция мандибуляторных желез, расположенных в голове и открывающихся в основании мандибул, не ясна. Известно, что они сильно развиты у некоторых муравьев, так что, видимо, имеют какое-то отношение к брачному поведению. Возможно также, что их выделениями метится добыча. У большинства![]() Myrmicinae и у Lasius выделения этих желез являются феромонами тревоги.
Myrmicinae и у Lasius выделения этих желез являются феромонами тревоги.

Рис. 30. Рабочие Monomorium (ориг.): а,б - M. dentigerum, в-д - M. barbatulum; в - тело в профиль; г - голова сверху; д - голова снизу; е,ж - M. gracillimum; з,и -
У ![]() Dolichoderinae феромон тревоги, имеющий характерный неприятный запах и выполняющий также репеллентные функции, выделяется анальной железой, открывающейся на вершине брюшка. У
Dolichoderinae феромон тревоги, имеющий характерный неприятный запах и выполняющий также репеллентные функции, выделяется анальной железой, открывающейся на вершине брюшка. У![]() Myrmicinae и
Myrmicinae и![]() Formicinae эта железа отсутствует. Также только у
Formicinae эта железа отсутствует. Также только у ![]() Dolichoderinae имеется железа Павана, расположенная в брюшке и открывающаяся между последним и предпоследним стернитами. Выделения её служат следовым феромоном. У
Dolichoderinae имеется железа Павана, расположенная в брюшке и открывающаяся между последним и предпоследним стернитами. Выделения её служат следовым феромоном. У![]() Ponerinae, Dorylinae и
Ponerinae, Dorylinae и![]() Formicinae следовые феромоны выделяются ректальными железами, открывающимися в заднюю кишку.
Formicinae следовые феромоны выделяются ректальными железами, открывающимися в заднюю кишку.
Очень разнообразны в разных группах муравьев функция желез ядовитого аппарата — ядовитой и железы Дюфура. Исходно смесь секретов этих желез образует яд, который насекомые вводят в жертву при помощи жала. Видимо, только эту функцию железы ядовитого аппарата имеют у![]() Ponerinae и Dorylinae. Хотя среди
Ponerinae и Dorylinae. Хотя среди

Myrmicinae и встречаются виды, способные жалить, например наш Myrmica, у многих представителей этого подсемейства жало используется не столько для защиты или нападения, сколько для нанесения пахучего следа на субстрат. В некоторых группах действующее начало следового феромона — секрет ядовитой железы (Monomorium, Tetramorium), в других — железы Дюфура (Atta, Pheidole, Solenopsis), в третьих — смесь этих веществ (Myrmica). Необычны среди![]() Myrmicinae представители рода Crematogaster, у которых следовые вещества выделяются тарзальными железами, расположенными в лапках.
Myrmicinae представители рода Crematogaster, у которых следовые вещества выделяются тарзальными железами, расположенными в лапках.
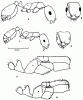
Рис. 32. Monomorium kusnezowi (а-в) и M. ruzskyi (голотип и паратип) (г-е) (ориг.): а-г - тело рабочего в профиль; б,д - голова рабочего сверху; в,е - грудь и стебелек самки в профиль.
У![]() Formicinae жало редуцировано и секреты ядовитых желез непосредственно изливаются на тело жертвы через ацидопору. Крайней степени эта способность достигает у Formica, у которых ядовитая железа гипертрофирована, мускулистый резервуар её занимает значительную часть брюшка, а яд (муравьиная кислота) может выбрызгиваться на расстояние нескольких десятков сантиметров. Секрет железы Дюфура служит, видимо, для маркировки добычи, а смесь этих веществ является также феромоном тревоги (Длусский, Чернышова,1974). У Lasius секрет железы Дюфура, имеющий запах герани, служит феромоном тревоги. Функция желез ядовитого аппарата
Formicinae жало редуцировано и секреты ядовитых желез непосредственно изливаются на тело жертвы через ацидопору. Крайней степени эта способность достигает у Formica, у которых ядовитая железа гипертрофирована, мускулистый резервуар её занимает значительную часть брюшка, а яд (муравьиная кислота) может выбрызгиваться на расстояние нескольких десятков сантиметров. Секрет железы Дюфура служит, видимо, для маркировки добычи, а смесь этих веществ является также феромоном тревоги (Длусский, Чернышова,1974). У Lasius секрет железы Дюфура, имеющий запах герани, служит феромоном тревоги. Функция желез ядовитого аппарата ![]() Dolichoderinae, также лишенных жала, не ясна, поскольку у этих муравьев защитную функцию выполняют выделения анальной железы.
Dolichoderinae, также лишенных жала, не ясна, поскольку у этих муравьев защитную функцию выполняют выделения анальной железы.
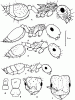
Рис. 33. Представители рода Crematogaster (ориг.): А - рабочий Cr. sorokini в профиль; Б - то же,
Особо следует сказать о метаплевральных железах муравьев, по-скольку их наличие — характерный признак надсемейства Formicoidea. Как показали Maschwitz et al. (1970), выделения этих желез обладают антибиотическим действием и предполагается, что основная их функция — дезинфекция гнезда. Возможно также, что они принимают участие в формировании «запаха гнезда».
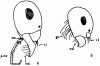
Рис. 34. Псаммофоры Cataglyphis setipes (А) и Messor intermedius (Б) (ориг.). Аммохеты: cl - клипеальные; gu - гулярные; md - мандибулярные; mn - ментальные; p.mx. - максиллярные шупики.
Промеры и индексы. При измерении муравьев используется несколько стандартных промеров и индексов. Измерения производят при помощи окуляр-микрометра. Использованы стандартные символы промеров в индексов, используемые в советской литературе по систематике муравьев:
ДМ — длина мезосомы от точки прикрепления головы до точки причленения петиолюса;
ДГ — длина головы от переднего края клипеуса до затылочного края, или, у видов с вогнутым затылочным краем, до условной линии, соединяющей затылочные углы;
ШГ — длина головы над глазами;
ИГ (головной индекс) — ШГ : ДГ;
ДС — длина скапуса от центра сочлененной головки до вершины; ИС (индекс скапуса) — ДС : ШГ;
ДГл — максимальная длина глаза; ИГл (глазной индекс) — ДГл : ШГ;
ДШ — длина щеки от нижнего края глаза до основания мандибулы.
Общая длина тела указывается приблизительно, т.к. измерить ее точно у муравьев практически невозможно из-за того, что сегменты брюшка втягиваются друг в друга. Размеры всегда даются в мм.

Рис. 6. Расчленение тела рабочих муравьев (ориг.):
А -
Б - Tapinoma erraticum (
В - Ponera coarctata
Г - Myrmica bessarabica
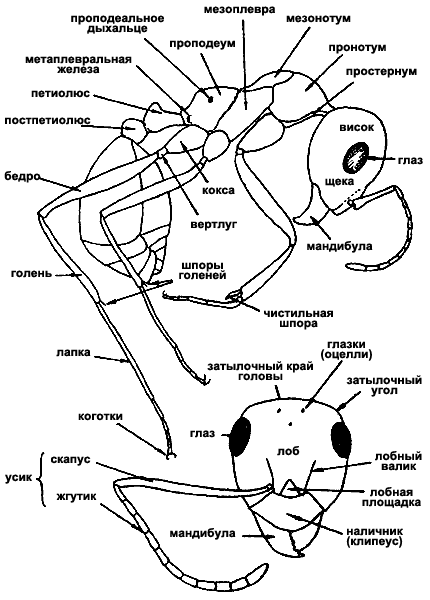
Рис. 9. Строение тела муравьев (ориг.):
вверху - рабочий
внизу - голова рабочего Formica subpilosa (сверху).
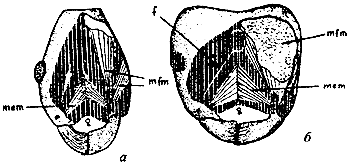
Рис. 7. Мускулатура головы Aphaenogaster raphidiiceps (а) и Messor aralocaspius (б) (Длусский, 1981): f - головная фурка; g - гулярный шов; mem - мышцы открывающие мандибулы; mfm - мышцы, сжимающие мандибулы.

Рис. 8. Тело самок сверху (а, в, д) и голова рабочих сверху (б, г, е) Diplorhoptrum(ориг.):
а, б -
в, Г - D. deserticolum;
д, е - D. celatum (д - голотип).


Рис. 11. Тело в профиль (А,Г,З), голова сверху (Б,Д,Е,И) и хетотаксия задней голени (В,Ж,К,Л) солдат и рабочего (Е) представителей подрода Tanaemyrmex рода Camponotus (ориг.): А,Б,В - C. turkestanus - Г-Ж - C. fedtschenkoi; 3,И,К - C. obliquipilosus; Л - C. bactrianus.
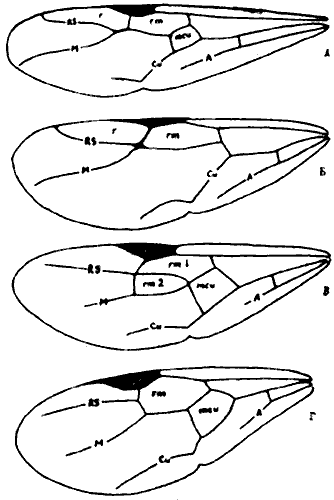
Рис. 49. Жилкование крыльев муравьев (ориг.): А - Formica subpilosa; Б - Camponotus obliquipilosus; В - Aphaenogaster gibbosa; Г - Crematogaster scutellaris. Свободные ветви жилок: A - анальной; Cu- кубитальной; M - медиальной; RS - радиус сектор. Ячейки: xmcu - медиокубитальная (дискоидальная); rm - радиомедиальные (кубитальные); r - радиальная.
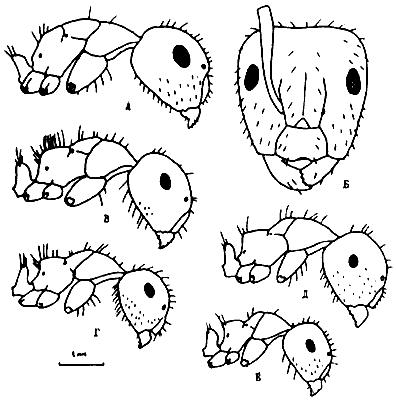
Рис. 12. Солдаты представителей подрода Myrmentoma рода Camponotus (ориг.);
А,Б - C. lameerei;
В - C. semirufus;
Г - C. interjectus;
Д - C. kopetdaghensis (паратип);
Е - C. lateralis.
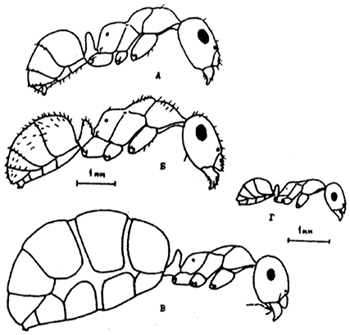
Рис. 3. Рабочие Formica и Proformica
(А,Б - ориг.; В,Г - по: Длусский (1981), с изменениями):
А - F. clara;
Б - F. subpilosa;
В - плерэргат P. epinotalis;
Г - мелкий рабочий P. epinotalis.
На рис. В и Г хетотаксия не изображена.
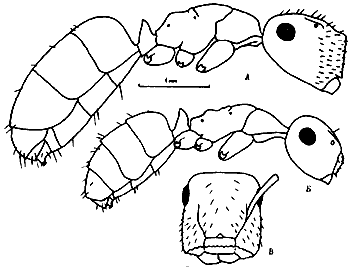

Рис. 14. Рабочий Polyrhachis simplex (Афганистан). Слева - тело в профиль, справа - грудь и петиолюс сверху (ориг.).
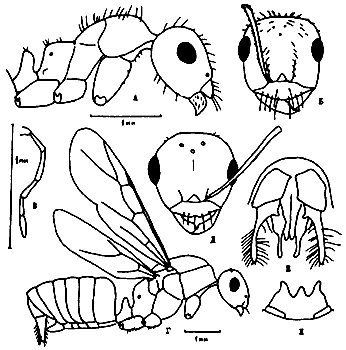
Рис. 15. Alloformica (ориг.): А,Б - A. obscurior sр.nov. (голотип) (А - тело в профиль, Б - голова сверху); В - максиллярный щупик рабочего A. aberrans; Г-Ж - самец A. aberrans (андртип) (Г - тело в профиль, Д - голова сверху, Е - гениталии сверху, Ж - субгенитальная пластинка снизу).
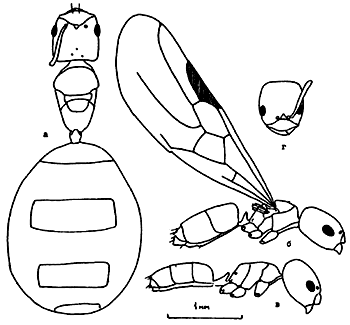
Рис. 4. Bothriomyrmex turcomenicus (ориг.):
а - старая (физогастрическая) самка;
б - молодая неоплодотворенная самка;
в - рабочий;
г - голова рабочего.
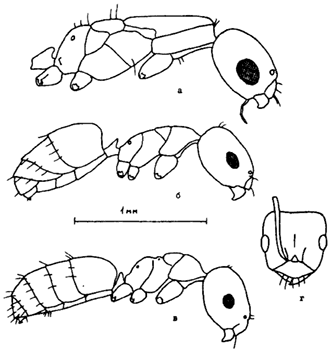
Рис. 16. Представители рода Plagiolepis (ориг.): а,б - Pl. pallescens (а - самка, б - рабочий); в,г - Pl. arnoldii sp.n. (голотип) (в - рабочий в профиль; г - голова того же экземпляра сверху).
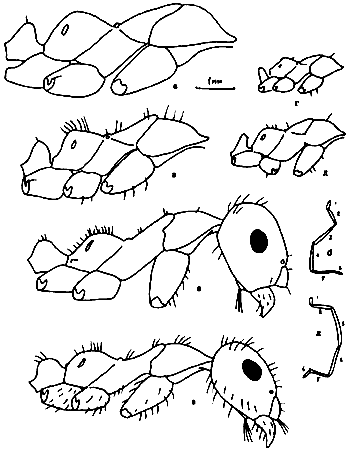
Рис. 17. Крупные виды Cataglyphis (ориг.): а,б - грудь в профиль и максиллярный щупик C. foreli (топотип); в - C. murgabica (паратип; Имам-Баба); г,д - крупный и мелкий рабочие
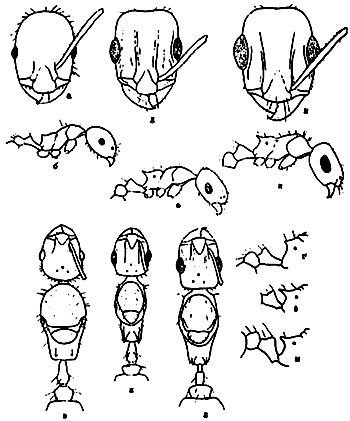
Рис. 18. Представители рода Temnothorax
(Длусский, Союнов, 1988). Голова рабочих сверху (а,д,и), тело рабочих в профиль (б,е,к), тело самок сверху (в,ж,л), стебелек и проподеум самок в профиль (г,з,м); а-г - T. recedens, д-з - T. semenovi, и-м - T. desertorum (голотип и паратип).
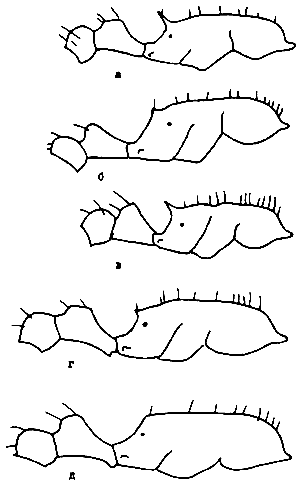
Рис. 19. Грудь и стебелек рабочих Leptothorax (ориг.):
а - L. unifasciatus (Копетдаг); б - L. melleus (паратип, Шугнан); в - L. parvulus (Кара-Кала); г - L. hasardaghi (паратип, Ел-Дере); д - L. anodontoides (паратип, Ай-Дере).

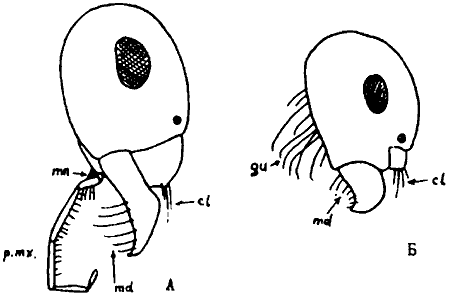
Рис. 34. Псаммофоры Cataglyphis setipes (А) и Messor intermedius (Б) (ориг.). Аммохеты: cl - клипеальные; gu - гулярные; md - мандибулярные; mn - ментальные; p.mx. - максиллярные шупики.
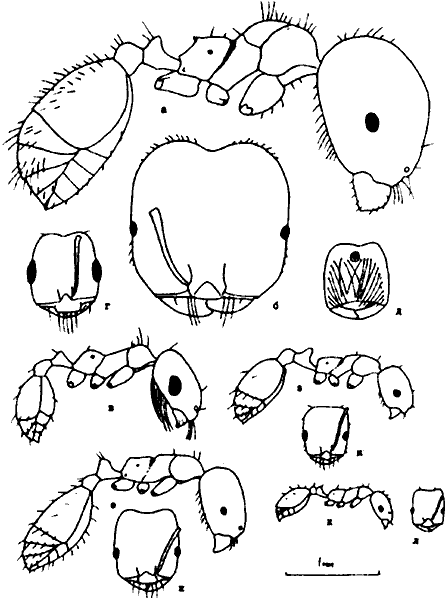
Рис. 30. Рабочие Monomorium (ориг.): а,б - M. dentigerum, в-д - M. barbatulum; в - тело в профиль; г - голова сверху; д - голова снизу; е,ж - M. gracillimum; з,и -
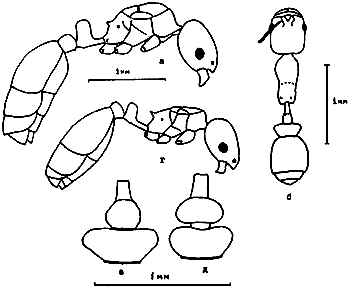
Рис. 21. Самки (а,г), петиолюс самок сверху (в,д) и рабочий (б) Cardiocondyla koshewnikovi (а-в) и C. elegans (г,д) (ориг.).
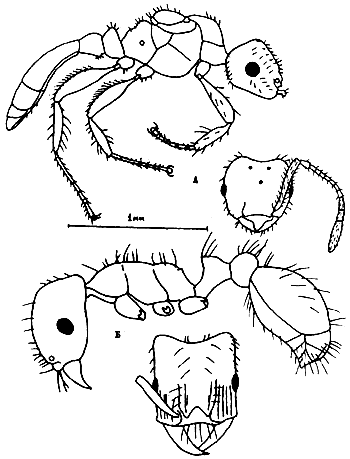
Рис. 22. Паразитические

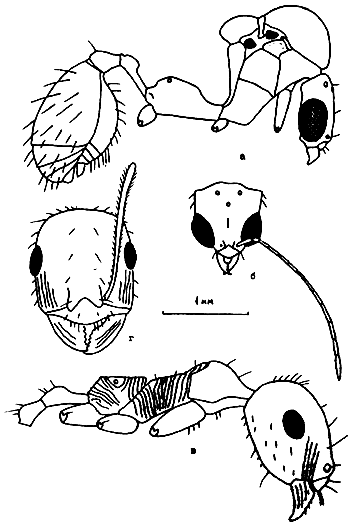
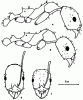
Рис. 24. Aphaenogaster messoroides sp. nov. (ориг.): а - самец (паратип); б - голова самца; в - рабочий (голотип); г - голова голотипа.
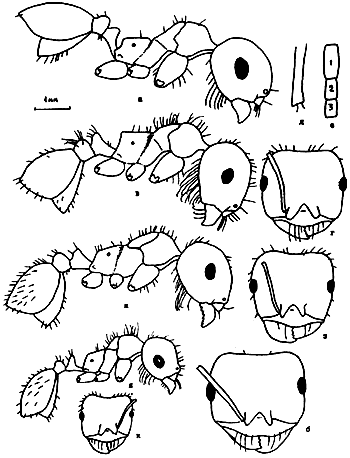
Рис. 25. Рабочие Messor из групп barbatulum (а–б) и aralocaspius (ж–к) (ориг.): а - M. intermedius, тело в профиль; б - он же, голова сверху; в - M. denticulatus, тело в профиль; г - он же, голова сверху; д - он же, основание скапуса; е - он же, 3 первые членика жгутика; ж - M. aralocaspius тело в профиль; з - он же, голова сверху; и - M. subgracilinodis (паратип), тело в профиль; к - он же, голова сверху.
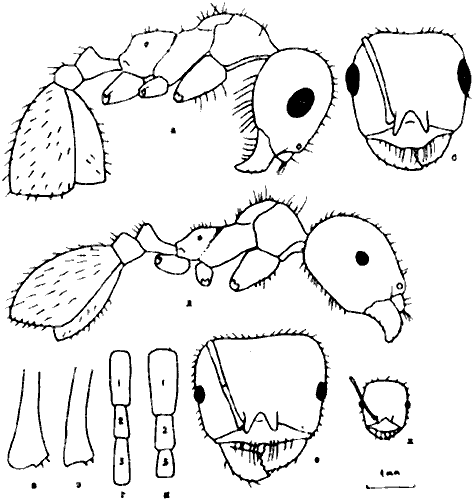
Рис. 26. Рабочие Messor из группы structor (ориг.); а–г –
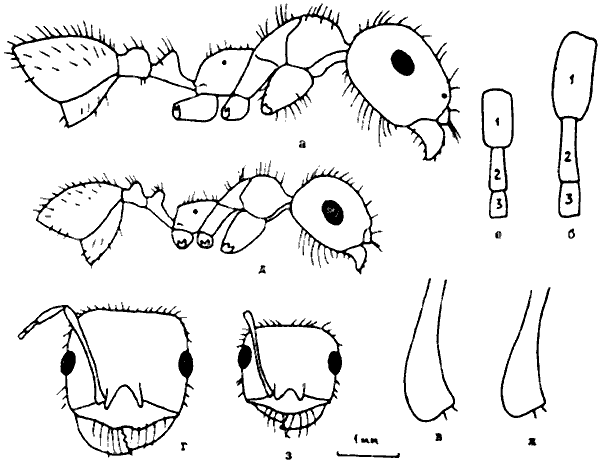
Рис. 27. Рабочие Messor из группы vicinus (ориг.): а-г - M. perantennatus; д-з - M. vicinus (а, д - тело в профиль; б,е - 3 первые членика жгутика; в,ж - основание скапуса; г,з - голова сверху).

Рис. 28. Рабочие Messor из группы excursionis (ориг.): а-в - M. excursionis; г-е - M. lamellicornis; ж-и - M. variabilis. (а,г,ж - тело в профиль; б,д,з - голова сверху; в,е,и -основание скапуса).

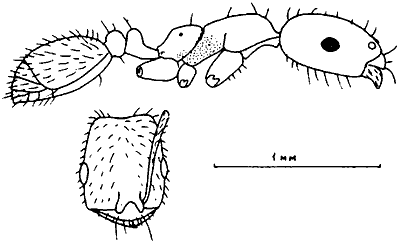
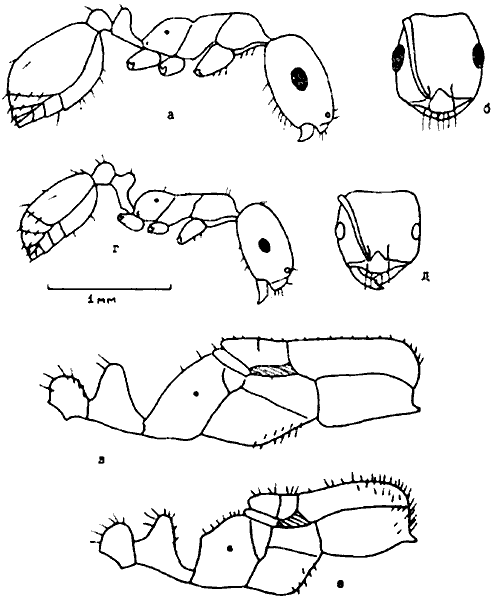
Рис. 32. Monomorium kusnezowi (а-в) и M. ruzskyi (голотип и паратип) (г-е) (ориг.): а-г - тело рабочего в профиль; б,д - голова рабочего сверху; в,е - грудь и стебелек самки в профиль.

Рис. 33. Представители рода Crematogaster (ориг.): А - рабочий Cr. sorokini в профиль; Б - то же,
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
грудная клетка, грудной отдел насекомых.
особый сегмент брюшка. У муравьев бывает двучлениковый (петиолюс + постпетиолюс) и одночлениковый (петиолюс - чешуйка). Количество члеников и форма стебелька очень важны для идентификации вида. Например у муравьев семейства Formicinae стебелек одночлениковый, а у Myrmicinae - двучлениковый.
 Lasius alienus (Foerster, 1850) — бледноногий муравей, бледный садовый муравей, полевой муравей, cornfield ant
Lasius alienus (Foerster, 1850) — бледноногий муравей, бледный садовый муравей, полевой муравей, cornfield antОдно из крупных подсемейств, насчитывает около 1,5 тыс. видов, относящихся к 44 родам. Широко распространены на всем земном шаре, довольно обычны в тропиках и занимают доминирующее положение в умеренных широтах.
Небольшое наиболее архаичное подсемейство муравьев. Распространено по всему земному шару, преимущественно в тропиках. Насчитывает более 230 видов.
Довольно примитивная группа муравьев по своему строению и поведению. Рабочие мономорфны, самки по размерам близки к рабочим.
Myrmicinae — самое большое подсемейство муравьев, включающее более 2 тыс. видов, относящихся более чем к 150 родам.
(от франц. facette грань) (сложные глаза), парный орган зрения, образован многочисленными отдельными глазками - омматидиями. Хорошо воспринимают движущиеся объекты, обеспечивают широкое поле зрения. Острота зрения и способность к восприятию формы предмета развиты слабо.
у муравьев имеется три основные касты: самцы, самки и рабочие (бесплодные модифицированные самки).
Всего 77 видов.
Всего 965 видов.
ксеробионт
(лат. mandibule — челюсть, от mando — жую, грызу), первая пара челюстей у ракообразных, многоножек и насекомых. Синоним: жвала.
первый членик усика, обычно намного длиннее члеников жгутика
Всего 71 вид.
это одна из форм социального паразитизма муравьев, при которой семья паразита живет отдельно от семьи хозяина, но периодически нападает на его гнезда и похищает куколок рабочих. Вышедшие из куколок «рабы» (или «помощники», по другой терминологии) выполняют свою обычную работу, но в гнезде паразита. Причина такого поведения «рабовладельцев» — частичная (у Raptiformica) или полная (у Polyergus, Rossomyrmex) редукция (утрата) касты рабочих.
Всего 6 видов.
Всего 26 видов.
 Diplorhoptrum fugax (Latreille, 1798) — муравей-вор
Diplorhoptrum fugax (Latreille, 1798) — муравей-воргигрофил
Старые названия: Solenopsis fugax, Solenopsis (fugax) orientalis,
Всего 172 вида.
Включает сокращенный Novomessor.
Всего 106 видов.
Всего насчитывает 549 видов.
Всего 14 видов.
 Lasius fuliginosus (Latreille, 1798) — пахучий муравей-древоточец, jet ants
Lasius fuliginosus (Latreille, 1798) — пахучий муравей-древоточец, jet antsчасть груди муравья, образоваванная слиянием заднегруди и первого сегмента брюшка.
 Messor structor (Latreille, 1798) — степной муравей-жнец
Messor structor (Latreille, 1798) — степной муравей-жнецСтарое название: Messor rufitarsis darianus Pisarski
часть груди муравья.
сегмент брюшка.
часть груди муравья. Состоит из: скутума и скутеллюма. (Извините, информация нуждается в дополнении/уточнении)
Всего 62 вида.
размещение (и тип) волосков на теле муравья, отличается большим постоянством и поэтому играет большую роль при определении вида.
гигрофил
сегмент брюшка.
Всего 65 видов.
Всего 26 видов.
 Camponotus truncatus (Spinola, 1808) — пробкоголовый муравей
Camponotus truncatus (Spinola, 1808) — пробкоголовый муравейполисахарид, образованный остатками аминосахара ацетилглюкозамина. Основной компонент наружного скелета (кутикулы) насекомых, ракообразных и других членистоногих. У грибов заменяет целлюлозу, с которой сходен по химическим и физическим свойствам и биологической роли.
Старое название: Polyrhachis simplex
короткая хитиновая трубочка, на конце брюшка, куда открывается кислая ядовитая железа.
венчик волосков, на конце брюшка, вокруг ацидопоры.
Всего 4 вида.
Всего 60 видов.
олигогинны
ксеробионт
Старые названия: Cataglyphis murgabica, Cataglyphis murgabicum, Cataglyphis foreli murgabica
 Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунок
Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунокгигрофил, галобионт
Старое название: Cataglyphis flavigastra
ксеробионт
Всего 319 видов.
Старое название: Leptothorax recedens
Старое название: Leptothorax semenovi
Всего 19 видов.
Старое название: Leptothorax unifasciatus
Старое название: Leptothorax balchashensis, Arnoldi, 1971
Старое название: Leptothorax parvulus
Старое название: Leptothorax anodontoides
специальное приспособление для переноски сухой почвы, распространенн у пустынных муравьев, образован длинными отстоящими волосками (аммохетами), часто имеющими форму латинской буквы J, расположен на нижней поверхности головы.
гигрофил
Всего 1 вид.
Всего 100 видов.
 Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелец
Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелецнадсемейство летающих насекомых отряда перепончатокрылых, родственное осам и муравьям. Существует около 20 тысяч видов пчёл. Распространены на всех континентах, кроме Антарктиды. Пчёлы питаются нектаром и пыльцой, используя нектар главным образом в качестве источника энергии, а пыльцу для получения белков и других питательных веществ.
органы осязания - отстоящие волоски, расположенные на последних члениках усика. При помощи отстоящих волосков муравьи также воспринимают движения воздуха и сотрясение субстрата.
(лат. receptio - прием) - органы и устройства, принимающие сигналы из внешней и внутренней среды организма и передающие их в центральную нервную систему.
гигрофил
живут колониями (пчелы, муравьи, термиты, некоторые осы). В состав колонии, напр. у термитов, входят половые особи (самцы и матки обеспечивают размножение), солдаты (защиту) и рабочие (собирание корма, строительство гнезда и т. д.). Солдаты и рабочие бесплодны; половые особи и солдаты сами не способны питаться; таким образом, члены колонии полностью зависят друг от друга. Разделение функций и сложные взаимодействия разных особей в колониях общественных насекомых обеспечиваются рядом особенностей в их строении и поведении; последнее регулируется с помощью выделяемых ими химических веществ (феромонов).
(греч. φέρω — «нести» + ορμόνη — «гормон») — летучие вещества, управляющие поведенческими реакциями, процессами развития, а также многими процессами, связанными с социальным поведением и размножением. Феромоны модифицируют поведение, физиологическое и эмоциональное состояние или метаболизм других особей того же вида.
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
 Monomorium pharaonis (Linnaeus, 1758) — pharaoh ant, фараонов муравей
Monomorium pharaonis (Linnaeus, 1758) — pharaoh ant, фараонов муравейВсего 415 видов
Всего 134 вида.
Всего 427 видов.
галобионт
Subspecies: Monomorium indicum kusnezowi Santschi, 1928
 Crematogaster schmidti (Mayr, 1853) — остробрюхие муравьи
Crematogaster schmidti (Mayr, 1853) — остробрюхие муравьиспецифический запах, служащий основой для разделения особей одного вида по принципу «свой - чужой»
гигрофил
часть гнезда рыжих лесных муравьев, занимаемая одной колонной.
Всего 24 вида.
рабочие с сильно раздутым брюшком; это каста, так называемых, "медовых бочек".
сильное вздутие/увеличение брюшка. Физогастрическая самка имеет гипертрофированные яичники, благодаря чему отличается огромной продуктивностью. Встречается у моногинных видов муравьев.






