






Муравьи практически не имеют конкурентов среди других беспозвоночных и поэтому выступают как относительно независимая группировка, состав которой определяют отношения конкуренции, мутуализма, паразитизма и т.п. между слагающими ее видами. Введенное нами (Длусский, 1981) для обозначения групп популяций муравьев, заселяющих один биогеоценоз, понятие коадаптивный комплекс подчеркивает их взаимную в процессе эволюции биоценоза приспособленность.
Анализ взаимной сопряженности видов муравьев в различных местообитаниях Туркменистана позволил выделить 3 комплекса, приуроченных к пустынным (I) (рис:40), предгорным и ксерофитным горным (II) и горно-степным (III) ландшафтам. Легко заметить (см. табл.1, табл.2, табл.3), что эти комплексы состоят из видов, имеющих существенно разное происхождение. Большинство видов, слагающих пустынный (I) комплекс, оформилось в процессе аридизации Средней Азии и соседних территорий. Большинство видов горно-степного (III) комплекса происходит из мезофильных областей Древнего Средиземноморья. Примечательно, что именно в этих регионах сформировались, соответственно принадлежности, все виды того и другого комплекса с наибольшим числом связей в плеядах, построенных по результатам вычисления коэффициентов взаимной сопряженности.
Итогом эволюции коадаптивного комплекса следует, очевидно, считать заполнение всех возможных для данной группы организмов экологических ниш, предоставляемых пространством заселяемого биогеоценоза. Для муравьев как основные параметры экологической ниши рассматриваются вид пищи, размеры её (коррелирующие с размерами тела), характер сезонной и суточной активности, используемые для сбора пищи ярусы. Распределение видов муравьев комплексов I—III по трофическим нишам показана в табл.4, табл.5, табл.6. Как показатель размера предпочитаемых пищевых объектов рассматриваются размерные классы самих муравьев, выраженные через сухой средний вес рабочего (Длусский, 1981): 1 — менее 0,1 мг; 2 — 0,1—0,25; 3 — 0,25—1,0; 4 — 1,0—2,5; 5 — 2,5—10,0; 6 — более 10,0 мг.
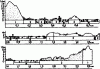
Рис. 40. Распределение муравьев по профилю грядово-такырного комплекса в Центральных Каракумах (Длусский, 1974): 1 - незакрепленные пески; 2 - закрепленные; 3 - такыровидные поверхности. Условные обозначения: ● - Cataglyphis pallida; ● - C. foreli; ● - C. takyrica; ● - Messor intermedius; ● - M. variabilis; ● - M. aralocaspius; ● - Pheidole pallidula; ● - Camponotus xerxes; ● - C. turkestanus; ● - Acantholepis semenovi.
Все виды пустынного комплекса собирают пищу в одном ярусе — герпетобях, за исключением декдробионта Crematogaster subdentata, В отличии от других представителей рода, у Messor variabilis фуражировка осуществляется одиночными сборщиками и, видимо, используются иные виды растений, так что между этим видом и доминантом Messor intermedius не возникает конкуренции, на что указывает наличие между ними достоверной связи. Другие 3 вида карпофагов 5-го размерного класса находятся в конкурентных отношениях, что и выражается в отсутствии корреляций в их распространении. В сумме виды комплекса используют 17 трофических ниш, что близко к теоретическому пределу. Незанятыми остаются ниши зоофагов 1-го размерного класса и зоофагов-дендробионтов 4-го размерного класса. Первую часто занимает Plagiolepis pallescens, а вторую — в пустынях в зарослями саксаула — Camponotus xerxes.
Виды адырного комплекса используют только 9 трофических ниш из 19 возможных. Поэтому в реальности пустующие ниши занимают виды пустынного комплекса в предгорьях или горно-степного — на сухих склонах гор. Общее число видов, выявляемых в коренных биотопах этого комплекса, остается в среднем тем же, что и в коренных биотопах других комплексов — 13—18.
Виды горно-степного комплекса активны преимущественно в теплое время года и в светлое время суток, а основные различия наблюдаются в использовании пространства по вертикали. Освоенное число ниш (13) близко к теоретически возможному.
Проведенный анализ позволяет сделать предположение, что эволюция пустынного и горно-степного комплексов практически завершена, тогда как адырный находится, скорее всего, в стадии становления.
Состав пустынного коадаптивного комплекса муравьев (зоогеографический аспект)
|
|||||||
Вид |
Число связей |
Регионы формирования видов |
|||||
1 |
2 |
3 |
4 |
5 |
6 |
||
| Cataglyphis emeryi | 11 |
+ |
|||||
| Monomorium barbatulum | 9 |
+ |
|||||
| Messor intermedius | 7 |
+ |
|||||
| Tetramorium schneideri | 7 |
+ |
|||||
| Tetramorium inerme | 7 |
+ |
|||||
| Messor variabilis | 6 |
+ |
|||||
| Messor subgracillinodis | 5 |
+ |
|||||
| Tetramorium reticuligerum | 5 |
+ |
|||||
| Camponotus turkestanus | 5 |
+ |
|||||
5 |
+ |
||||||
| Cerapachys desertorum | 5 |
+ |
|||||
| Cataglyphis setipes | 4 |
+ |
|||||
| Cataglyphis cinnamomea | 4 |
+ |
|||||
| Crematogaster sorokini | 3 |
+ |
|||||
| Messor denticulatus | 2 |
+ |
|||||
| Cardiocondyla koshewnikovi | 2 |
+ |
|||||
2 |
+ |
||||||
| Camponotus turkestanicus | 2 |
+ |
|||||
| Messor aralocaspius | 1 |
+ |
|||||
| Crematogaster subdentata | 1 |
+ |
|||||
| Итого: | 93 |
14 |
4 |
1 |
1 |
||
Примечание. Здесь и в табл.2,3: 1 - пустыни Туранской низменности; 2 - пустыни Индостана; 3 - горы и низкогорья Хорасана; 4 - степи Евразии; 5 - аридные горы Средиземноморья; 6 - широколиственные леса Средиземноморья. |
|||||||
Состав адырного или полусаванного коадаптивного комплекса муравьев (зоогеографический аспект)
|
|||||||
Вид |
Число связей |
Регионы формирования видов |
|||||
1 |
2 |
3 |
4 |
5 |
6 |
||
| Monomorium ruzskyi | 4 |
+ |
|||||
| Tetramorium karakalense | 3 |
+ |
|||||
| Acantholepis semenovi | 3 |
+ |
|||||
| Cataglyphis nodus | 3 |
+ |
|||||
| Plagiolepis pallescens | 2 |
+ |
|||||
| Camponotus bactrianus | 2 |
+ |
|||||
| Crematogaster bogojawlenskii | 1 |
+ |
|||||
| Tetramorium armatum | 1 |
+ |
|||||
| Messor intermedius | 1 |
+ |
|||||
| Итого: | 20 |
2 |
1 |
3 |
3 |
||
Состав горно-степного коадаптивного комплекса муравьев (зоогеографический аспект)
|
|||||||
Вид |
Число связей |
Регионы формирования видов |
|||||
1 |
2 |
3 |
4 |
5 |
6 |
||
| Aphaenogaster gibbosa | 12 |
+ |
|||||
| Camponotus aethiops | 9 |
+ |
|||||
8 |
+ |
||||||
| Leptothorax parvulus | 7 |
+ |
|||||
7 |
+ |
||||||
6 |
+ |
||||||
| Messor turcmenochorassanicus | 5 |
+ |
|||||
| Formica clara | 5 |
+ |
|||||
5 |
+ |
||||||
| Leptothorax elderensis | 4 |
+ |
|||||
| Leptothorax hasardaghi | 4 |
+ |
|||||
| Camponotus kopetdaghensis | 4 |
+ |
|||||
| Diplorhptrum fugax | 1 |
+ |
|||||
| Temnothorax recedens | 1 |
+ |
|||||
| Итого: | 77 |
3 |
1 |
3 |
7 |
||
Распределение муравьев пустынного комплекса по трофическим нишам
|
|||
Параметры классификации |
Вид |
||
Вид пищи |
Размерный класс |
Период активности |
|
| Растительная | 1 |
круглогодично,днем | Cardiocondyla koshewnikovi |
2 |
круглогодично, зимой — днем, другие сезоны — утром и вечером |
Monomorium barbatulum | |
2 |
апрель—июнь и сентябрь—октябрь, 19—24ч | Tetramorium inerme | |
2 |
апрель—май, ночью | Tetramorium reticuligerum | |
2 |
июнь—сентябрь, утром и вечером | ||
3 |
круглогодично, круглосуточно | Tetramorium schneideri | |
4 |
с осени до весны, днем | Messor surgacillinodis | |
5 |
зимой—весной — днем летом и осенью — ночью. |
Messor intermedius Messor denticulatus Messor aralocaspius Messor variabilis |
|
| Животная | 2 |
май—сентябрь, ночью | Cerapachys desertorum |
3 |
февраль—май, первая половина дня | Cataglyphis emeryi | |
3 |
май—сентябрь, вторая половина дня | Cataglyphis cinnamomea | |
4 |
круглогодично, днем | ||
4 |
круглогодично, ночью | Camponotus turkestanus | |
5 |
кроме зимы, днем | Cataglyphis setipes | |
5 |
кроме зимы, ночью | Camponotus turkestanicus | |
| Падь тлей и мелкие насекомые |
3 |
кроме зимы, днем | Crematogaster sorokini Crematogaster subdentata |
Распределение муравьев адырного комплекса по трофическим нишам
|
|||
Параметры классификации |
Вид |
||
Вид пищи |
Размерный класс |
Период активности |
|
| Растительная | 2 |
кроме зимы, круглосуточно | Monomorium ruzskyi |
2 |
апрель—июнь, сентябрь—ноябрь — круглосуточно | Tetramorium karakalense | |
2 |
апрель—июнь, сентябрь, ночью | Tetramorium armatum | |
5 |
круглогодично: зимой—весной — днем, летом—осенью — ночью | Messor intermedius | |
| Животная | 1 |
кроме зимы, в сумерки и ночью | Plagiolepis pallescens |
2 |
кроме зимы, днем | Acantholepis semenovi | |
5 |
кроме зимы, днем | Cataglyphis nodus | |
5 |
кроме зимы, ночью | Camponotus bactrianus | |
| Падь тлей и мелкие насекомые |
2 |
кроме зимы, круглосуточно | Crematogaster bogojawlenskii |
Распределение муравьев горно-степного комплекса по трофическим нишам
|
|||
Параметры классификации |
Вид |
||
Вид пищи |
Размерный класс |
Предпочитаемый ярус |
|
| Растительная | 2 |
Герпетобий | |
2 |
Подстилка | Temnothorax recedens | |
5 |
Герпетобий | Messor turcmenochorassanicus | |
| Растительная и животная |
4 |
Подстилка и герпетобий | Aphaenogaster gibbosa |
| Животная | 1 |
Почва | |
2 |
Почва | ||
2 |
Подстилка | Leptothorax elderensis | |
2 |
Все ярусы | Leptothorax parvulus | |
3 |
Все ярусы | Leptothorax hasardaghi | |
4 |
Все ярусы | Camponotus aethiops | |
| Падь тлей и мелкие насекомые |
2 |
Все ярусы | |
3 |
Древостой | ||
4 |
Герпетобий, травостой | Camponotus kopetdaghensis | |
4 |
Герпетобий, древостой | Formica clara | |

Рис. 40. Распределение муравьев по профилю грядово-такырного комплекса в Центральных Каракумах (Длусский, 1974): 1 - незакрепленные пески; 2 - закрепленные; 3 - такыровидные поверхности. Условные обозначения: ● - Cataglyphis pallida; ● - C. foreli; ● - C. takyrica; ● - Messor intermedius; ● - M. variabilis; ● - M. aralocaspius; ● - Pheidole pallidula; ● - Camponotus xerxes; ● - C. turkestanus; ● - Acantholepis semenovi.
животные, не имеющие позвоночника: простейшие, губки, кишечнополостные, черви, моллюски, членистоногие, иглокожие и др.; примерно 3/4 видов всех животных.
(лат. mutuus - взаимный) - форма взаимодействия организмов, выгодная и обязательная для объединяющихся организмов и популяций, например, многие животные не могут существовать без микроорганизмов в их кишечнике.
совокупность особей одного вида, длительно занимающая определенное пространство и воспроизводящая себя в течение большого числа поколений. В современной биологии популяция рассматривается как элементарная единица процесса эволюции, способная реагировать на изменения среды перестройкой своего генофонда.
эволюционно сложившаяся, пространственно ограниченная, длительно самоподдерживающаяся, однородная экологическая система, в которой функционально взаимосвязаны живые организмы и окружающая их абиотическая среда.
необратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.
(от греч. oikos дом, жилище, местопребывание и ...логия), наука об отношениях живых организмов и образуемых ими сообществ между собой и с окружающей средой. Термин «экология» предложен в 1866 Э. Геккелем. Объектами экологии могут быть популяции организмов, виды, сообщества, экосистемы и биосфера в целом.
ксеробионт
ксеробионт
Старое название: Pheidole arenarum Ruzsky
Старые названия: Acantholepis semenovi, Acantholepis transcaspiensis
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
организм, питающийся семенами растений.
организм, питающийся исключительно другими животными
организм, обитающий в древесном ярусе растительности.
олигогинны
(гр. topos - место) - естественное жизненное пространство определенного биоценоза, совместно с которым он образует экосистему.
ксеробионт
Старое название: Proformica deserta
Всего 106 видов.
Всего 415 видов
 Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунок
Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунокгигрофил, галобионт
Старое название: Cataglyphis flavigastra
ксеробионт
Старое название: Cataglyphis cinnamomea
гигрофил
гигрофил
 Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушитель
Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушительСтарое название: Monomorium gracillimum
 Tetramorium caespitum (Linnaeus, 1758) — дерновый муравей, pavement ant
Tetramorium caespitum (Linnaeus, 1758) — дерновый муравей, pavement antСтарое название: T. indocile
Старое название: Leptothorax parvulus
 Lasius alienus (Foerster, 1850) — бледноногий муравей, бледный садовый муравей, полевой муравей, cornfield ant
Lasius alienus (Foerster, 1850) — бледноногий муравей, бледный садовый муравей, полевой муравей, cornfield ant Crematogaster schmidti (Mayr, 1853) — остробрюхие муравьи
Crematogaster schmidti (Mayr, 1853) — остробрюхие муравьи Lasius flavus (Fabricius, 1782) — желтый луговой (садовый, земляной) муравей
Lasius flavus (Fabricius, 1782) — желтый луговой (садовый, земляной) муравейВсего 19 видов.
Старое название: Leptothorax recedens
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
 Diplorhoptrum fugax (Latreille, 1798) — муравей-вор
Diplorhoptrum fugax (Latreille, 1798) — муравей-воргигрофил
Старые названия: Solenopsis fugax, Solenopsis (fugax) orientalis,






