






Участок, на котором муравьи из одной семьи собирают пищу носит название кормового. У наиболее высокоорганизованных муравьев, семьи которых насчитывают более 100 тыс. особой (Formica s. str., Dendrolasius, Liometopum и др.), кормовой участок полностью охраняется от особей своего вида из других семей или от других видов со сходной биологией. В этом случае говорят об охраняемой территории. Хотя такие виды в фауне Туркменистана отсутствуют, элементы территориальности имеются и у наших видов. Во многих случаях охраняется не весь кормовой участок, а только пригнездовая зона. Такое поведение характерно для Cataglyphis и Acanthoiepis. У муравьев-жнецов (Messor) кормовые участки семей перекрываются в пространстве, но не во времени. Эти муравьи каждый день заново формируют кормовой участок и охраняют его, но одни и те же куртины эфемеров могут в разные дни посещаться муравьями из разных семей (см. рис:39).
У примитивных хищников и некрофагов, например у Ponera coarctata, Cataglyphis emeryi или Alloformica aberrans, рабочие-фуражиры ведут себя на кормовом участке совершенно независимо. Однако в большинстве случаев у муравьев существует те или иные способы повышения эффективности использования кормового участка. Простейшая форма такого взаимодействия — неспецифическая активация фуражиров. Муравей, обнаруживший пищу (разведчик), возвращается в гнездо и там вызывает возбуждение других муравьев. Каждый из возбужденных фуражиров направляется туда, где он обычно добывает корм. В результате, число муравьев на кормовом участке резко возрастает, что повышает вероятность случайного нахождения того же или другого источника корма. Такое поведение характерно, например, для фаэтончика Cataglyphis setipes. В середине дня, когда становится жарко, муравьи-жнецы покидают свои кормовые участки. Во время фуражировки они одновременно выносят из гнезда и выбрасывают трупы муравьев, погибших за ночь в гнезде, но в это время фаэтончики не могут проникнуть на кормовой участок, т.к. его охраняют жнецы. После ухода жнецов в гнездо, трупы становятся доступными и стоит 1—2 фаэтончикам вернуться в гнездо с трупами жнецов, как из гнезда выбегает десяток или более муравьев, каждый из которых направляется искать трупы на свои индивидуальные кормовые участки.
Наиболее эффективное средство организации групповой фуражировки — мобилизация. Под ней понимается комплекс действий разведчика, который приводит к тому, что другие муравьи (пассивные фуражиры) точно выходят к месту, где разведчиком был обнаружен корм. Способы мобилизации у разных видов муравьев могут сильно отличаться. Простейший способ — киноптическая мобилизация, был описан Г.М.Длусским 1981 а) у пустынного бегунка Cataglyphis foreli при охоте на термитов. Стоит рабочему этого вида вернуться в гнездо с термитом, как из него в результате неспецифической активации выходят 10—30 муравьев, которые, однако, никуда не уходят, а стоят около входа и наблюдают за поведением разведчика. После второго возвращения разведчика часть из мобилизованных муравьев уходит в том направлении, откуда тот вернулся. Таким образом, передается направление к источнику пищи, но не указывается расстояние до него.
Фактически развитием этого способа является своеобразная мобилизация пустынных муравьев-жнецов (Захаров, 1972; Длусский, 1931а) Организацию групповой фуражировки у них осуществляют активные фуражиры (разведчики), составляющие всего 3% от их общего числа. Обычно они работают как одиночные сборщики на своих индивидуальных участках площадью 0,2—0,4 м² и все вместе собирают информацию о ситуации на всем потенциальном кормовом участке. В пустыне у Messor aralocaspius и M. denticulatus при интенсивной заготовке семян выделяются два периода активности — с рассвета до середины дня и со второй половины дня, когда температура на поверхности опускается до 35—39°, до наступления темноты. При этом муравьи каждый раз заново формируют фуражировочные дороги и таким образом последовательно эксплуатируют разные куртины эфемеров в пределах их кормового участка. Когда на последнем создаются благоприятные условия для фуражировки, из гнезда появляется большое число (150—300) пассивных фуражиров, которые скапливаются у входа. Из этой массы выбегают несколько активных фуражиров и начинают совершать короткие пробежки в том направлении, где они во время предыдущего выхода нашли много созревших семян. Пассивные фуражиры устремляются за ними. Формируется тропа из муравьев, которые начинают медленно (со скоростью 15—20 см/с) двигаться в заданном направлении. После этого активные фуражиры уже не участвуют в процессе, но тропа постепенно доходит до поля созревших эфемеров, где пассивные фуражиры начинают сбор семян. Такой способ мобилизации, названный нами дистанционным наведением, не дает большой точности, но она и не нужна, т.к. потенциальный источник корма занимает большую площадку. В то же время он позволяет за короткое время мобилизовать несколько сотен и даже тысяч фуражиров.
Другие способы мобилизации связаны с точным наведением пассивных фуражиров на источник пищи, простейшим из них — движение тандемом. У Cardiocondyla и Leptothorax разведчик, вернувшись в гнездо, активизирует одного рабочего. Когда он вновь направляется к корму, мобилизованный муравей идет за ним следом, постоянно прикасаясь усиками к брюшку и таким образом приходит к корму. Обратно они возвращаются самостоятельно и приводят еще двух новых муравьев. У Myrmica, Tetramorium, Monomorium barbatulum и некоторых Camponotus наблюдается привод группы. Разведчик, вернувшись в гнездо после обнаружения корма, активизирует других муравьев при помощи сложного комплекса действий (пробежки, подрагивания телом, прикосновение усиками и лапками), получившего название осцилляторной рекрутирующей демонстрации (ОРД). Активизированные рабочие выходят из гнезда и выстраиваются за разведчиком цепочкой,которую он и приводит к корму.
Многие муравьи используют для мобилизация пахучий след. Разведчик на пути от корма к гнезду оставляет пахучий след выделениями специальных желез. У![]() Formicinae и
Formicinae и![]() Ponerinae — это ректальные, у
Ponerinae — это ректальные, у ![]() Dolichoderinae железа Павана, расположенная на нижней стороне брюшка, у Crematogaster — тарзальные железы и у других
Dolichoderinae железа Павана, расположенная на нижней стороне брюшка, у Crematogaster — тарзальные железы и у других![]() Myrmicinae — ядовитая железа (Monomorium, Tetramorium) или железа Дюфура (Pheidole, Solenopsis). У Pheidole pallidula пахучий след сам по себе без дополнительной активизации служит для других муравьев сигналом к мобилизации. Каждый фуражир, нашедший след, оставленный разведчиком, начинает двигаться по нему в направлений от гнезда и находит корм. У Lasius, (Tapinoma, Crematogaster и многих Monomorium реагируют на след только фуражиры, предварительно активизированные разведчиком при помощи ОРД.
Myrmicinae — ядовитая железа (Monomorium, Tetramorium) или железа Дюфура (Pheidole, Solenopsis). У Pheidole pallidula пахучий след сам по себе без дополнительной активизации служит для других муравьев сигналом к мобилизации. Каждый фуражир, нашедший след, оставленный разведчиком, начинает двигаться по нему в направлений от гнезда и находит корм. У Lasius, (Tapinoma, Crematogaster и многих Monomorium реагируют на след только фуражиры, предварительно активизированные разведчиком при помощи ОРД.
Каждый из этих способов мобилизации имеет свои преимущества и недостатки и зависит от того, какое место в биоценозе занимает вид. Дистанционное наведение позволяет быстро мобилизовать большое число рабочих, но при этом точность наведения довольно низка, так что способ хорош для мобилизации на большие кормовые площади, но непригоден для мобилизации на «точечный» источник пищи (труп, небольшие группы растений и т.д.). Использование пахучего следа позволяет за короткое время привести большое число пассивных фуражиров на «точечный» источник, но этот способ очень инерциален и после исчезновения пищи муравьи еще долго продолжают ходить по пахучей тропе. Таким образом, он лучше всего обеспечивает использование «точечных», но довольно крупных источников пищи (например, трупы крупных насекомых или мелких позвоночных животных). Мобилизация путем привода группы менее эффективна, но зато группы муравьев могут быстро переключаться с одного источника на другой, а семья в целом — использовать одновременно несколько источников пищи.
Другой путь повышения эффективности использования кормового участка связан с совершенствованием структурной организации кормового участка и основан на вторичном делении его. В простейшем случае, например, у Cataglyphis, каждый фуражир имеет индивидуальный поисковый участок, на котором ведет поиск и сбор пищи. Каждый индивидуальный участок при этом представляет собой сектор в пределах кормового участка семьи.
У Acantholepis semenovi каждая семья имеет охраняемую территорию. Между территориями разных семей имеются нейтральные зоны, шириной 20—40 см. В пределах территории можно выделить 3 зоны. В пригнездовой зоне с радиусом 20—30 см действуют в основном муравьи-строители, выносящие почву из гнезда. В ближней зоне охоты (до 2—3 м от гнезда) охота и транспортировка добычи ведутся коллективно. При появлении в этой зоне источника пищи разведчики производят в гнезде неспецифическую активизацию, в результате которой плотность муравьев увеличивается. В дальней зоне охоты (до 8—10 м) активные фуражиры (сборщики трупов и одновременно охотники) ведут поиск на индивидуальных участках площадью около 0,25 м. Участки отдельных особей перекрываются. При обнаружении крупной добычи активизация в гнезде не производится, но рабочий, нашедший добычу, которую не может самостоятельно умертвить или доставить в гнездо, начинает возбужденно бегать вокруг нее и тем самым привлекает внимание других муравьев, которые приходят к нему на помощь, В дальней зоне охоты работают и сборщики пади, которые движутся по сходным территориям, так что образуются потоки.
Еще сложнее устроена территория у некоторых видов Formica, в частности у рыжих лесных муравьев (группа ![]() Formica rufa). У них имеются колонии — система связанных между собой гнезд, в каждом из которых обитает отдельная семья. На общей территории колонии имеется сложная система постоянных дорог. Поскольку представители этой группы в Туркменистане не встречаются, мы не останавливаемся подробно на этом вопросе, отсылая читателя к работам А.А.Захарова (1972, 1978).
Formica rufa). У них имеются колонии — система связанных между собой гнезд, в каждом из которых обитает отдельная семья. На общей территории колонии имеется сложная система постоянных дорог. Поскольку представители этой группы в Туркменистане не встречаются, мы не останавливаемся подробно на этом вопросе, отсылая читателя к работам А.А.Захарова (1972, 1978).
У пустынных Monomorium описана система использования кормового участка, включающая специфичное гнездостроение и мобилизацию. Так, у Monomorium kusnezovi и ![]() M. gracillimum от гнезда в разных направлениях отходят длинные поверхностные галереи с многими входами. С каждым входом связана небольшая более или менее постоянная группа рабочих, фуражирующая вокруг входа. У Monomorium barbatulum имеется многосекционное гнездо, секции которого удалены друг от друга на расстояние около 1м и соединены подземными галереями. Каждая секция содержит постоянную группу из 100—300 рабочих, фуражирующих вблизи гнезда. Хотя радиус поиска и среднее расстояние мобилизации у этих муравьев невелики (среднее для Monomorium barbatulum 0,74 м, M. kusnezovi — 1,09 м,
M. gracillimum от гнезда в разных направлениях отходят длинные поверхностные галереи с многими входами. С каждым входом связана небольшая более или менее постоянная группа рабочих, фуражирующая вокруг входа. У Monomorium barbatulum имеется многосекционное гнездо, секции которого удалены друг от друга на расстояние около 1м и соединены подземными галереями. Каждая секция содержит постоянную группу из 100—300 рабочих, фуражирующих вблизи гнезда. Хотя радиус поиска и среднее расстояние мобилизации у этих муравьев невелики (среднее для Monomorium barbatulum 0,74 м, M. kusnezovi — 1,09 м, ![]() M. gracillimum — 0,61 м), семья в целом может эксплуатировать кормовой участок более 10 м².
M. gracillimum — 0,61 м), семья в целом может эксплуатировать кормовой участок более 10 м².
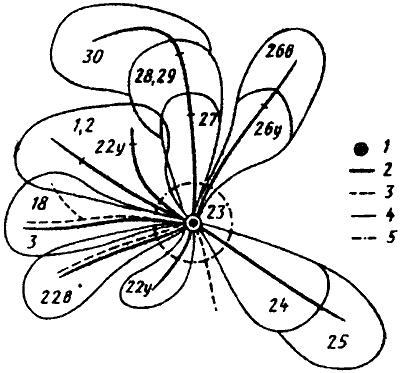
Рис. 39. Использование кормового участка фуражирами одной семьи Messor aralocaspius (Захаров, 1972): 1 - гнездо; 2 - дороги с 22 апреля по 3 мая 1970 г.; 3 - дорога до 16 апреля; 4 - граница сбора семян; 5 - пригнездовая территория. Цифрами обозначены даты фуражировки по секторам. Индексы: у - утренняя, в - вечерняя фуражировки.
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
Всего 100 видов.
Всего 14 видов.
участок, используемый муравьями для добычи пищи.
участок, охраняемый семьей (колонной) от посещения чужими особями своего или близких видов.
Всего 65 видов.
Всего 106 видов.
(зоонекрофаг), организм, питающийся мертвыми животными, гетеротроф
ксеробионт
Старое название: Proformica deserta
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
ксеробионт
индивидуальное или групповое привлечение пассивных фуражиров активными фуражирами для конкретных полей в конкретном месте.
ксеробионт
отряд общественных насекомых. Общины, разделенные на касты, состоят из крылатых и бескрылых особей. Строят подземные и наземные (до 15 м высотой) гнезда (термитники). Ок. 2600 видов, главным образом в тропиках; в России 2 вида: один в районе Сочи, второй, вероятно, завезенный из Японии или Китая, во Владивостоке. Многоядны. Разрушают древесину, портят бумагу, кожу, сельскохозяйственные продукты и др. Термиты, обитающие в почве, полезны как почвообразователи.
ксеробионт
гигрофил
Всего 38 видов.
Всего 19 видов.
Всего 415 видов
Всего 965 видов.
Одно из крупных подсемейств, насчитывает около 1,5 тыс. видов, относящихся к 44 родам. Широко распространены на всем земном шаре, довольно обычны в тропиках и занимают доминирующее положение в умеренных широтах.
Довольно примитивная группа муравьев по своему строению и поведению. Рабочие мономорфны, самки по размерам близки к рабочим.
Небольшое наиболее архаичное подсемейство муравьев. Распространено по всему земному шару, преимущественно в тропиках. Насчитывает более 230 видов.
Всего 427 видов.
Myrmicinae — самое большое подсемейство муравьев, включающее более 2 тыс. видов, относящихся более чем к 150 родам.
Всего насчитывает 549 видов.
Всего 134 вида.
Старое название: Pheidole arenarum Ruzsky
Всего 62 вида.
эволюционно сложившаяся, пространственно ограниченная, длительно самоподдерживающаяся, однородная экологическая система, в которой функционально взаимосвязаны живые организмы и окружающая их абиотическая среда.
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
часть гнезда рыжих лесных муравьев, занимаемая одной колонной.
Старые названия: Acantholepis semenovi, Acantholepis transcaspiensis
пространство между охраняемыми территориями семей или колонн не посещаемое фуражирами из конкурирующих семей (колонн).
сладкая густая жидкость, выделяемая тлями, червецами и другими насекомыми, питающимися соком растений. Медоносная пчела собирает падь с листьев и перерабатывает в падевый мед.
 Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравейфункциональное образование, состоящее из материнской и дочерних семей, поддерживающих регулярные родственные отношения
галобионт
Subspecies: Monomorium indicum kusnezowi Santschi, 1928
 Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушитель
Monomorium destructor (Jerdon, 1851) — изящный муравей, муравей-разрушительСтарое название: Monomorium gracillimum
гнездо, состоящее из повторяющихся стереотипных блоков (секций), каждый из которых включает все видоспецифичные элементы (ствол, камеры и т. д.)






