






Даже в травяном ярусе имеется несколько зон, к которым могут специально приспособиться общественные насекомые; это корни, поверхность почвы и травостой. Смене грасслендов формацией кустарников и лесной формацией могут препятствовать разнообразные травоядные животные; или же грассленды могут сохраняться благодаря таким факторам, как отсутствие воды и непрерывные неблагоприятные вмешательства со стороны человека (особенно пожары, в частности в саванне). Мы рассмотрим обе эти ситуации.
Solenopsis представлен в Северной Америке комплексом из шести видов; два из них были интродуцированы в южные районы США и распространились там. Один из них, S. richteri («черный огненный муравей»), прибыл сюда в 1918 г. из Аргентины и Уругвая, а второй — S. invicta («красный огненный муравей») — в 1933—1945 гг. из Мату-Гросу (Бразилия). Ни тот, ни другой не изменился по сравнению с исходными южноамериканскими популяциями, и скрещивание их происходит редко (Lofgren et al., 1975). В настоящее время распространение S. richteri ограничено северными частями штатов Миссисипи и Алабама, тогда как S. invicta широко распространился от Техаса до Северной Каролины. Обычно в семье бывает одна самка, а иногда две, и известна по меньшей мере одна высокополигинная семья. Предполагается, что S. invicta, отличающийся агрессивностью, в большинстве районов вытеснил S. richteri благодаря своей большей способности выслеживать и уничтожать самок S. richteri, собирающихся основать семью. S. invicta подавил также местный вид S. xyloni, так как оба они обитали в грасслендах; однако четвертый вид, S. geminata, сохранился в лесных местностях, хотя первоначально он тоже обитал в грассленде. Тем не менее S. xyloni не вполне вытеснен, так как Baroni Urbani и Kannowski (1974) нашли его в грассленде вместе с S. invicta; однако он живет здесь лишь на участках, находящихся в тени деревьев, на которых он фуражирует, избегая таким образом встреч с S. invicta. Даже если расставлялись приманки с целью увеличить контакты между этими видами, они все же избегали друг друга и дело никогда не доходило до прямых схваток. Семьи S. invicta агрегированы, что вызывает удивление, так как этот вид обычно моногинен и имеет четко ограниченные территории; можно предположить, что какие-то условия среды варьируют здесь случайным образом. Wilson (1978) провел сравнительное изучение видов S. invicta и S. geminata. Он измерял время, затрачиваемое рабочими особями разных размерных групп на выполнение всего репертуара своих функций. Оба вида полиморфны, но крупные рабочие особи у S. geminata намного больше, чем у S. invicta. У обоих видов самые мелкие рабочие остаются в гнезде, тогда как крупные переносят частицы почвы и пищи, соответствующие им по величине. У S. invicta крупные рабочие особи пережевывают семена и запасают жиры, а у S. geminata они заняты исключительно дроблением семян; они даже не нападают на добычу. В природе S. invicta более всеядны, чем S. geminata, которые поедают семена многих типов.
Myrmica — голарктический род со многими видами (рис:88). Низкий температурный оптимум (20°C) делает его приспособленным к жизни в областях с прохладным летом. Для развития потенций репродуктивной самки требуется по меньшей мере 6-недельное воздействие температуры около 10°C в зимнее время; поэтому на юге этот род встречается только в горах. Пара видов, обычная для невозделываемых травянисто-вересковых пустошей в Европе, — это M. ruginodis и M. scabrinodis, хорошо различимые по величине и морфологии; Elmes (1978b, 1980а) провел биометрическое исследование этих двух видов. Они различаются по микроместообитаниям, что можно связать с особенностями поведения. M. scabrinodis устраивает гнезда в низкой травянистой растительности или под лишайниками между группами растений вереска; при устройстве подземной части гнезда, разделенной на мелкие камеры, эти муравьи широко используют глину. Такие гнезда превосходно поглощают солнечное тепло, устойчивы к высыханию и очень эффективны в качестве укреплений, когда этот вид захватывает гнезда M. ruginodis, занимая одну камеру за другой. M. scabrinodis охотится у поверхности почвы, между основаниями стеблей, а компактное строение его тела дает ему возможность пробираться сквозь густую траву. Таким образом, благодаря особенностям своего образа жизни он может обитать в самых жарких участках злаково-вересковой ассоциации, на пятнах густой низкой травы. M. ruginodis устраивает гнезда в высокой траве и злаках, часто вблизи кустов или даже в рыхлом мхе. Надземные постройки его гнезда менее массивны; это скорее оболочка или полог, покрывающий обширную поверхностную камеру, разделенную несколькими тонкими перегородками. Такое устройство гнезда позволяет им подниматься к солнцу и поглощать тепло лучей, пробивающихся сквозь высокую траву, а если тень становится слишком густой — перебираться на новое место; однако гнездо не обладает устойчивостью к высыханию и не защищено от нападения врагов. M. ruginodis кормится в травяном ярусе выше, чем M. scabrinodis; он взбирается на кусты, в чем ему очень помогают длинные ноги и тонкие торчащие вперед усики. Подобно M. scabrinodis, он использует в пищу разнообразных мелких животных, как живых, так и мертвых, а также падь, но фуражирует в верхних частях растений.
Итак, экология этих двух видов достаточно различна, чтобы они могли сосуществовать в местообитании одного типа; у них выработались структурные и поведенческие различия, предотвращающие столкновения между ними. Конфликты возникают только на участках с густой низкой травой, где селится M. scabrinodis; здесь он не только защищает свою собственность, но и активно отбрасывает M. ruginodis. M. scabrinodis может осадить гнездо M. ruginodis, и тогда всей семье последнего в конце концов приходится уйти на новое место, а M. scabrinodis занимает гнездо и перестраивает его на свой лад; если же M. ruginodis не уходит, то M. scabrinodis вторгается в гнездо, завоевывая одну камеру за другой. Сухая погода помогает M. scabrinodis, так как его собственные гнезда защищают их обитателей от обезвоживания. Таким образом, контакт между этими двумя видами ослаблен благодаря тому, что они фуражируют и гнездятся в разных ярусах общего местообитания. В то время как один вид (M. scabrinodis) может полностью доминировать над другим на теплых сухих участках и лучше использует их, другой (M. ruginodis) способен выжить за пределами этих зон, где на него не оказывает давления M. scabrinodis, но где подходящий микроклимат распределен пятнами. Элмер (Elmer, 1978а) изучал Myrmica в вересковой пустоши на юге Англии и подтвердил эти различия в распространении между M. ruginodis и M. scabrinodis, отметив, что второй из них вытесняет первого в «... более бедные и неоднородные вересковые районы», тогда как сам он доминирует в «...более однородных участках с развитой дерновиной Calluna». Элмер высказывает также мнение, что микрогинная форма M. ruginodis «...дает этому виду возможность полностью использовать неоднородные районы...»
Дальше на восток, где климат теплее, а почва более нейтральная, эта пара комплементарных видов встречается еще с двумя видами — ![]() M. rubra и M. sabuleti. Из рис:88 сразу видно, что каждому виду северной пары соответствует свой вид в южной паре:
M. rubra и M. sabuleti. Из рис:88 сразу видно, что каждому виду северной пары соответствует свой вид в южной паре: ![]() M. rubra перекрывается с M. ruginodis, а M. sabuleti — с M. scabrinodis. Это могло бы затруднить их идентификацию, если бы к таксономическим критериям не добавлялись особенности поведения, на которые натуралисты научились полагаться. Хотя вид
M. rubra перекрывается с M. ruginodis, а M. sabuleti — с M. scabrinodis. Это могло бы затруднить их идентификацию, если бы к таксономическим критериям не добавлялись особенности поведения, на которые натуралисты научились полагаться. Хотя вид ![]() M. rubra по своему телосложению, подобно M. ruginodis, приспособлен к фуражированию в траве, он живет на участках с нейтральной почвой, т. е. в местах, обычно используемых для выпаса скота, но не для интенсивного сельского хозяйства. Здесь растет чертополох, а на нем живут тли; упавшие со стен камни предоставляют муравьям готовые гнезда, которым не угрожает затаптывание. В грасслендах на известняках
M. rubra по своему телосложению, подобно M. ruginodis, приспособлен к фуражированию в траве, он живет на участках с нейтральной почвой, т. е. в местах, обычно используемых для выпаса скота, но не для интенсивного сельского хозяйства. Здесь растет чертополох, а на нем живут тли; упавшие со стен камни предоставляют муравьям готовые гнезда, которым не угрожает затаптывание. В грасслендах на известняках ![]() M. rubra распространен пятнами, главным образом в тенистых местах, где ему не угрожает конкуренция со стороны
M. rubra распространен пятнами, главным образом в тенистых местах, где ему не угрожает конкуренция со стороны ![]() Lasius flavus, которому нужно больше солнца. Несмотря на это,
Lasius flavus, которому нужно больше солнца. Несмотря на это, ![]() M. rubra, вероятно, живет в более теплых местах, чем M. ruginodis, который вытесняется в разреженные кустарники и редколесье (Elmes, Wardlaw, 1982). Вид M. sabuleti, сходный по телосложению с M. scabrinodis, фуражирует вблизи поверхности земли и живет в грасслендах, которые либо выпасаются (если почва нейтральная), либо растительность на них распределена пятнами (если почва кислая); в обоих случаях муравьи могут устраивать здесь гнезда в местах, полностью освещаемых солнцем, или строить небольшие холмики. M. scabrinodis, судя по его более южному географическому распространению, больше нуждается в тепле, чем M. sabuleti.
M. rubra, вероятно, живет в более теплых местах, чем M. ruginodis, который вытесняется в разреженные кустарники и редколесье (Elmes, Wardlaw, 1982). Вид M. sabuleti, сходный по телосложению с M. scabrinodis, фуражирует вблизи поверхности земли и живет в грасслендах, которые либо выпасаются (если почва нейтральная), либо растительность на них распределена пятнами (если почва кислая); в обоих случаях муравьи могут устраивать здесь гнезда в местах, полностью освещаемых солнцем, или строить небольшие холмики. M. scabrinodis, судя по его более южному географическому распространению, больше нуждается в тепле, чем M. sabuleti.
Описанные два вида сосуществуют в грасслендах с кислой почвой (pH 5,2) на юго-западе Англии. M. sabuleti лучше чувствует себя в местах с более редкой растительностью, тогда как M. scabrinodis обитает на участках, заросших высокими злаками; различие в высоте травы на тех и других участках может быть незначительным. В грасслендах с известковой почвой (pH 7,8) примерно на той же широте встречается только M. sabuleti, но здесь на выеденных и вытоптанных местах этот вид менее обилен. Растительность вокруг гнезд этого муравья отличается от остальной местной растительности; лучшие гнезда расположены на открытых местах, и обычно вблизи от них растет Hieracium pilosella (сложноцветные) (Elmes, Wardlaw, 1982b). De Vroey (1979) показал, что в Бельгии M. sabuleti и M. scabrinodis пространственно разобщены, а Doncaster (1981), исследовавший эти же два вида на уэльском морском острове Рамси, установил, что M. sabuleti — более эвритопный вид и способен жить при изменении двух главных факторов среды — экспозиции склона и дренирования — в гораздо более широком диапазоне, чем M. scabrinodis. По третьему, относительно менее важному фактору — высоте, а также наличию орляка и приморской растительности, M. sabuleti отходит от M. scabrinodis и характеризуется как обитатель низких скалистых мест (края утесов) с тонким слоем почвы и лишайниками, хорошо освещаемых солнцем и теплых. M. sabuleti обитает на большей высоте, на холмах, где прохладнее и где он тесно ассоциируется с ![]() Lasius flavus.
Lasius flavus.
Эти четыре вида Myrmica — M. ruginodis, ![]() M. rubra, M. Scabrinodis и M. sabuleti — делятся, таким образом, на две пары, в каждую из которых входит один южный и один северный представитель, один обитатель низкой и один — высокой травы. Если выращивать перезимовавших личинок при определенной температуре, то они окукливаются в определенной последовательности: если время до окукливания M. ruginodis принять за единицу, то для
M. rubra, M. Scabrinodis и M. sabuleti — делятся, таким образом, на две пары, в каждую из которых входит один южный и один северный представитель, один обитатель низкой и один — высокой травы. Если выращивать перезимовавших личинок при определенной температуре, то они окукливаются в определенной последовательности: если время до окукливания M. ruginodis принять за единицу, то для ![]() M. rubra оно составит 1,2, для M. scabrinodis 1,4 и для M. sabuleti 1,5 (Elmes, Wardlaw 1982c,d). У всех видов Q10 = 3. Средняя температура на глубине 5 см в местообитании M. ruginodis равна 13,1°С,
M. rubra оно составит 1,2, для M. scabrinodis 1,4 и для M. sabuleti 1,5 (Elmes, Wardlaw 1982c,d). У всех видов Q10 = 3. Средняя температура на глубине 5 см в местообитании M. ruginodis равна 13,1°С, ![]() M. rubra — 13,9°С, M. scabrinodis — 14,3°С и M. sabuleti — 16,0°С, т. е. они располагаются в той же самой последовательности.
M. rubra — 13,9°С, M. scabrinodis — 14,3°С и M. sabuleti — 16,0°С, т. е. они располагаются в той же самой последовательности.
В роде Messor, возможно, наблюдается такая же картина. На юго-западе Франции встречаются два вида: моногинный ![]() M. capitatus с крупными рабочими и очень большими семьями, у которого новую семью закладывают, вероятно, одиночные самки, и полигинный
M. capitatus с крупными рабочими и очень большими семьями, у которого новую семью закладывают, вероятно, одиночные самки, и полигинный ![]() M. structor, в семьях которого бывает до 17 самок (Delage, 1968).
M. structor, в семьях которого бывает до 17 самок (Delage, 1968). ![]() M. capitatus приурочен к сухим склонам с известковой почвой, густо поросшим злаками, где он процветает (вместе с Pheidole pallidula), тогда как гораздо менее специализированный
M. capitatus приурочен к сухим склонам с известковой почвой, густо поросшим злаками, где он процветает (вместе с Pheidole pallidula), тогда как гораздо менее специализированный ![]() M. structor может занимать более разнообразные местообитания, в том числе долины и известняковые плато, где он устраивает глубокие разветвленные гнезда, избегая тем самым обезвоживания во время засухи; он часто живет вокруг мест, занятых
M. structor может занимать более разнообразные местообитания, в том числе долины и известняковые плато, где он устраивает глубокие разветвленные гнезда, избегая тем самым обезвоживания во время засухи; он часто живет вокруг мест, занятых ![]() M. capitatus. В сущности, можно задать вопрос: не является ли пара видов
M. capitatus. В сущности, можно задать вопрос: не является ли пара видов ![]() M. structor и
M. structor и ![]() M. capitatus экологическим эквивалентом пары Myrmica ruginodis и M. scabrinodis? Существует много видов Messor, но особенно интересны два вида, изученные в саванне Ламто (Берег Слоновой Кости) (Levieux, 1979; Levieux, Diomande, 1978а): M. galla живет в местах, поросших редкой и низкой травой, а M. regalis — в высокой траве; это различие, снова заставляющее вспомнить Myrmica, приводит к тому, что они собирают семена разных видов злаков. Как будто для того, чтобы встреча между ними стала еще менее вероятной, они фуражируют в разное время дня. При возвращении фуражировочных колонн Messor galla в гнездо 60—70% рабочих особей бывают нагружены, и они заняты сборами в саванне круглый год. Муравьи M. regalis фуражируют в одиночку и спорадически, если не считать сухого сезона, когда корм имеется в изобилии, и нагруженными возвращаются только 40—50% рабочих.
M. capitatus экологическим эквивалентом пары Myrmica ruginodis и M. scabrinodis? Существует много видов Messor, но особенно интересны два вида, изученные в саванне Ламто (Берег Слоновой Кости) (Levieux, 1979; Levieux, Diomande, 1978а): M. galla живет в местах, поросших редкой и низкой травой, а M. regalis — в высокой траве; это различие, снова заставляющее вспомнить Myrmica, приводит к тому, что они собирают семена разных видов злаков. Как будто для того, чтобы встреча между ними стала еще менее вероятной, они фуражируют в разное время дня. При возвращении фуражировочных колонн Messor galla в гнездо 60—70% рабочих особей бывают нагружены, и они заняты сборами в саванне круглый год. Муравьи M. regalis фуражируют в одиночку и спорадически, если не считать сухого сезона, когда корм имеется в изобилии, и нагруженными возвращаются только 40—50% рабочих.
Различны и гнезда этих двух видов: у M. galla гнездо диффузное (и семья, вероятно, полигинная), а M. regalis строит купол примерно с тремя отверстиями по 10 см в диаметре каждое. Таким образом, эти виды, принадлежащие к одному роду, различаются по местообитаниям, пище, времени активности, способу сбора пищи и строению гнезд — более чем достаточно для экологической дифференциации.
Crematogaster — весьма многообразный род![]() Myrmicinae — представлен в саванне Ламто двумя видами: один из них, Cr. heliophila, живет на верхушках деревьев, а другой, Cr. impressa, — в кустах, у поверхности почвы (Delage-Darchen, 1971). Оба вида очень сходны морфологически. Cr. heliophila строит гнезда из картона; Cr. impressa обычно обитает внутри веточек, строит также непрочные картонные гнезда. Оба питаются соками растений и продуктами жизнедеятельности щитовок, но насекомых они поедают разных, поскольку охотятся в разных растительных ярусах. В семьях Cr. heliophila одна самка, они очень агрессивны и могут занимать несколько соседних деревьев. Семьи Cr. impressa легко распадаются на фрагменты, особенно после пожаров, и каждый фрагмент принимает по нескольку новых самок, хотя этот вид не всегда бывает полигинным и число маток может периодически уменьшаться, как у ос. Распределение по ярусам усиливается, по крайней мере отчасти, враждебностью рабочих особей Cr. heliophila, хотя возможно, что некоторое влияние оказывает и выбор местообитания самкой. Явное сходство с Myrmica ruginodis и M. scabrinodis состоит здесь в том, что Cr. heliophila занимает теплый лесной полог и строит прочные гнезда, вытесняя муравьев Cr. impressa в более прохладные, менее стабильные слои, в которых они выживают благодаря большей подвижности, достигаемой за счет снижения расхода энергии на постройку гнезда.
Myrmicinae — представлен в саванне Ламто двумя видами: один из них, Cr. heliophila, живет на верхушках деревьев, а другой, Cr. impressa, — в кустах, у поверхности почвы (Delage-Darchen, 1971). Оба вида очень сходны морфологически. Cr. heliophila строит гнезда из картона; Cr. impressa обычно обитает внутри веточек, строит также непрочные картонные гнезда. Оба питаются соками растений и продуктами жизнедеятельности щитовок, но насекомых они поедают разных, поскольку охотятся в разных растительных ярусах. В семьях Cr. heliophila одна самка, они очень агрессивны и могут занимать несколько соседних деревьев. Семьи Cr. impressa легко распадаются на фрагменты, особенно после пожаров, и каждый фрагмент принимает по нескольку новых самок, хотя этот вид не всегда бывает полигинным и число маток может периодически уменьшаться, как у ос. Распределение по ярусам усиливается, по крайней мере отчасти, враждебностью рабочих особей Cr. heliophila, хотя возможно, что некоторое влияние оказывает и выбор местообитания самкой. Явное сходство с Myrmica ruginodis и M. scabrinodis состоит здесь в том, что Cr. heliophila занимает теплый лесной полог и строит прочные гнезда, вытесняя муравьев Cr. impressa в более прохладные, менее стабильные слои, в которых они выживают благодаря большей подвижности, достигаемой за счет снижения расхода энергии на постройку гнезда.
Род Lasius ![]() (Formicinae) изучался в грасслендах умеренного пояса Европы. Pontin (1961a,b) исследовал взаимоотношения между черным наземным
(Formicinae) изучался в грасслендах умеренного пояса Европы. Pontin (1961a,b) исследовал взаимоотношения между черным наземным ![]() L. niger и желтым подземным
L. niger и желтым подземным ![]() L. flavus в Англии; он обнаружил, что первый вид поедает второй и оказывает пагубное влияние на продукцию его половых особей. Однако масштабы уничтожения
L. flavus в Англии; он обнаружил, что первый вид поедает второй и оказывает пагубное влияние на продукцию его половых особей. Однако масштабы уничтожения ![]() L. flavus муравьями
L. flavus муравьями ![]() L. niger невелики из-за того, что последние — плохие строители гнезд и могут жить только там, где плоские камни предоставляют им готовое природное гнездо. Эти два вида тоже кормятся в разных растительных ярусах:
L. niger невелики из-за того, что последние — плохие строители гнезд и могут жить только там, где плоские камни предоставляют им готовое природное гнездо. Эти два вида тоже кормятся в разных растительных ярусах: ![]() L. flavus питается почти исключительно почвенными членистоногими и выращивает на корнях растений тлей многих видов, тогда как
L. flavus питается почти исключительно почвенными членистоногими и выращивает на корнях растений тлей многих видов, тогда как ![]() L. niger взбирается на травинки и кусты, собирает «дикую» падь, охотится и поедает падаль. На любом пастбище с грубой травой, где есть плоские камни и немного кустарников, эти два вида распределяются по разным ярусам и сосуществуют, но при высоких плотностях
L. niger взбирается на травинки и кусты, собирает «дикую» падь, охотится и поедает падаль. На любом пастбище с грубой травой, где есть плоские камни и немного кустарников, эти два вида распределяются по разным ярусам и сосуществуют, но при высоких плотностях ![]() L. niger частично уничтожает
L. niger частично уничтожает ![]() L. flavus.
L. flavus.
Эта пара видов — важные члены сообщества на о. Вадден (Нидерланды). ![]() Lasius niger первым проникает на только что заселенные растениями ложбины между дюнами, а
Lasius niger первым проникает на только что заселенные растениями ложбины между дюнами, а ![]() L. flavus и другие виды, обитающие под землей, следуют за ним только тогда, когда выше уровня грунтовых вод образуется достаточный слой почвы, который позволяет избегать вредного засоления и делает возможным рост таких злаков, как Festuca rubra (Boomsma, van Loon, 1982). Затем
L. flavus и другие виды, обитающие под землей, следуют за ним только тогда, когда выше уровня грунтовых вод образуется достаточный слой почвы, который позволяет избегать вредного засоления и делает возможным рост таких злаков, как Festuca rubra (Boomsma, van Loon, 1982). Затем ![]() L. flavus может значительно превзойти по численности
L. flavus может значительно превзойти по численности ![]() L. niger и оказать сильное отрицательное влияние на размеры его рабочих особей и производство репродуктивных самок, но не самцов. Эти данные служат дополнением к результатам исследований Pontin в Англии (Boomsma et al., 1982).
L. niger и оказать сильное отрицательное влияние на размеры его рабочих особей и производство репродуктивных самок, но не самцов. Эти данные служат дополнением к результатам исследований Pontin в Англии (Boomsma et al., 1982).
![]() Lasius alienus гораздо ближе к
Lasius alienus гораздо ближе к ![]() L. niger как по строению, так и по поведению. Он встречается в более южных районах, где ему не грозит опасность со стороны более крупных и более агрессивных рабочих
L. niger как по строению, так и по поведению. Он встречается в более южных районах, где ему не грозит опасность со стороны более крупных и более агрессивных рабочих ![]() L. niger, но на юге Англии, где обитают оба вида,
L. niger, но на юге Англии, где обитают оба вида, ![]() L. alienus может лишь избегать
L. alienus может лишь избегать ![]() L. niger и, чтобы получить необходимое ему тепло, выбирает сухие, открытые склоны, хорошо освещаемые солнцем (Brian, 1964; Brian et al., 1976). Здесь он устраивает свои подземные
гнезда и прокладывает фуражировочные тоннели, что обеспечивает ему защиту от ветра, дождя и пожаров, периодически возникающих на пустошах. Его основную пищу составляют почвенные членистоногие, особенно после пожара, но, кроме того, он тесно связан с тлей Aphis ulicis, обитающей на стелющихся по земле побегах Ulex minor. В отсутствие фуражиров
L. niger и, чтобы получить необходимое ему тепло, выбирает сухие, открытые склоны, хорошо освещаемые солнцем (Brian, 1964; Brian et al., 1976). Здесь он устраивает свои подземные
гнезда и прокладывает фуражировочные тоннели, что обеспечивает ему защиту от ветра, дождя и пожаров, периодически возникающих на пустошах. Его основную пищу составляют почвенные членистоногие, особенно после пожара, но, кроме того, он тесно связан с тлей Aphis ulicis, обитающей на стелющихся по земле побегах Ulex minor. В отсутствие фуражиров ![]() L. niger рабочие особи
L. niger рабочие особи ![]() L. alienus выходят на поверхность и взбираются на папоротники и кустарники в поисках нектара и пади; рабочие
L. alienus выходят на поверхность и взбираются на папоротники и кустарники в поисках нектара и пади; рабочие ![]() L. niger могут скопляться вокруг выходных отверстий гнезд
L. niger могут скопляться вокруг выходных отверстий гнезд ![]() L. alienus и нападать на муравьев, пытающихся выйти наружу. Брачный лёт у
L. alienus и нападать на муравьев, пытающихся выйти наружу. Брачный лёт у ![]() L. niger происходит в июле-сентябре, а у
L. niger происходит в июле-сентябре, а у ![]() L. alienus в сентябре-октябре, так что существует некоторая опасность гибридизации между ними; Pearson (1982а) путем изозимного анализа установил, что такая гибридизация действительно происходит. Он показал также, что у
L. alienus в сентябре-октябре, так что существует некоторая опасность гибридизации между ними; Pearson (1982а) путем изозимного анализа установил, что такая гибридизация действительно происходит. Он показал также, что у ![]() L. alienus возможен хромосомный полиморфизм — встречаются диплоидные числа хромосом 28 и 30.
L. alienus возможен хромосомный полиморфизм — встречаются диплоидные числа хромосом 28 и 30.
Самки ![]() Lasius niger, по-видимому, избегают сухих пустошей, служащих излюбленным местообитанием
Lasius niger, по-видимому, избегают сухих пустошей, служащих излюбленным местообитанием ![]() L. alienus; вероятно, для
L. alienus; вероятно, для ![]() L. niger в июле там слишком жарко и сухо, хотя они и селятся на голой земле в садах (Pontin, 1960), подобно самкам американского L. neoniger, которые избегают лесов, и если поместить их туда после того, как они сбросили крылья, отказываются рыть гнезда (Wilson, Hunt, 1966). В южной части Англии Elmes (1971) перемещал
L. niger в июле там слишком жарко и сухо, хотя они и селятся на голой земле в садах (Pontin, 1960), подобно самкам американского L. neoniger, которые избегают лесов, и если поместить их туда после того, как они сбросили крылья, отказываются рыть гнезда (Wilson, Hunt, 1966). В южной части Англии Elmes (1971) перемещал ![]() L. niger в местообитания
L. niger в местообитания ![]() L. alienus, и
L. alienus, и ![]() L. niger здесь постепенно уничтожался. Таким образом, эти виды созданы для различных местообитаний, но иногда перекрываются; там, где это происходит, приуроченность к разным ярусам растительности дополняется прямым поведенческим взаимодействием, которое снижает продукцию половых особей у обоих видов, а тем самым и их приспособленность. Поэтому умение молодых самок различать местообитания приобретает большое значение.
L. niger здесь постепенно уничтожался. Таким образом, эти виды созданы для различных местообитаний, но иногда перекрываются; там, где это происходит, приуроченность к разным ярусам растительности дополняется прямым поведенческим взаимодействием, которое снижает продукцию половых особей у обоих видов, а тем самым и их приспособленность. Поэтому умение молодых самок различать местообитания приобретает большое значение.
На острове Рамси в грасслендах, выедаемых кроликами, Lasius neoniger селится вдали от побережья, у старых стен, большие камни которых предоставляют ему идеальные места для гнезд (Doncaster, 1981). ![]() L. alienus, напротив, встречается почти исключительно на побережье и живет на вершинах утесов в подземных тоннелях. Свои диффузные гнезда он строит под очень низкой травой или под лишайниками; они открыты ветрам, но также и солнечным лучам, и в начале лета подвержены периодическому иссушению. Применяя метод главных компонент, Doncaster показал, что эти два вида совершенно не перекрываются в тех двух измерениях, от которых зависит примерно 89% их дисперсии: L. neoniger приурочен к укрытым, умеренно влажным участкам, а
L. alienus, напротив, встречается почти исключительно на побережье и живет на вершинах утесов в подземных тоннелях. Свои диффузные гнезда он строит под очень низкой травой или под лишайниками; они открыты ветрам, но также и солнечным лучам, и в начале лета подвержены периодическому иссушению. Применяя метод главных компонент, Doncaster показал, что эти два вида совершенно не перекрываются в тех двух измерениях, от которых зависит примерно 89% их дисперсии: L. neoniger приурочен к укрытым, умеренно влажным участкам, а ![]() L. alienus — к открытым и сухим. Иными словами, местообитание
L. alienus — к открытым и сухим. Иными словами, местообитание ![]() L. alienus на острове примерно такое же, как в Англии; а
L. alienus на острове примерно такое же, как в Англии; а ![]() L. niger, вероятно, селится в садах и на вересковых пустошах, т. е. в местах, заменяющих его болотное местообитание в Англии.
L. niger, вероятно, селится в садах и на вересковых пустошах, т. е. в местах, заменяющих его болотное местообитание в Англии. ![]() L. flavus — наиболее широко распространенный из всех видов, живущих на острове Рамси; его способность самому строить себе холмики и фуражировать под землей позволяет ему жить в тех местах, где не очень сильное выедание травы кроликами снижает ее рост; однако при высокой плотности кроликов они могут разрушать муравейники, раскапывая землю когтями. Методом главных компонент
L. flavus — наиболее широко распространенный из всех видов, живущих на острове Рамси; его способность самому строить себе холмики и фуражировать под землей позволяет ему жить в тех местах, где не очень сильное выедание травы кроликами снижает ее рост; однако при высокой плотности кроликов они могут разрушать муравейники, раскапывая землю когтями. Методом главных компонент ![]() L. flavus можно четко отличить от
L. flavus можно четко отличить от ![]() L. niger и
L. niger и ![]() L. alienus, введя третью компоненту, отражающую высоту, для которой характерны орляк (Pteridium aquilinum) и прибрежная растительность. Таким образом, эти три вида хорошо дифференцированы по своим экологическим особенностям в трехмерном пространстве компонент со следующими расстояниями: 3,14 между
L. alienus, введя третью компоненту, отражающую высоту, для которой характерны орляк (Pteridium aquilinum) и прибрежная растительность. Таким образом, эти три вида хорошо дифференцированы по своим экологическим особенностям в трехмерном пространстве компонент со следующими расстояниями: 3,14 между ![]() L. alienus и
L. alienus и ![]() L. niger, 2,61 между
L. niger, 2,61 между ![]() L. alienus и
L. alienus и ![]() L. flavus и 2,66 между
L. flavus и 2,66 между ![]() L. flavus и
L. flavus и ![]() L. niger.
L. niger.
Camponotus представляет собой, вероятно, самый обычный род муравьев (Wilson, 1976d), распространенный от тропиков до умеренных поясов. На одной лишь территории Берега Слоновой Кости (Западная Африка) описано 50 видов; экологию некоторых из них изучал Levieux (1976). C. acvapimensis — обычнейший вид, обитающий в почве саванны; его биомасса составляет 0,52 г/м² в перегнойно-карбонатных почвах, 0,47 г/м² в ферралитах и 0,2 г/м² в гидроморфных почвах (по сухому весу). Если самки после брачного лёта садятся на занятую территорию, рабочие особи уничтожают их. Другой вид, C. solon, у которого рабочие достигают в длину более 1 см, устраивает гнезда в стволах высоких деревьев, далеко отстоящих одно от другого (1—2 на гектар), и питается выделениями растений, собирая их в пологе деревьев, удаленных от гнезда на расстояние до 60 м.
Единовременный кормовой участок занимает несколько квадратных метров листвы, но за год муравьи используют несколько деревьев. Разведчики поднимают свои семьи с наступлением ночи, из гнезда выходит толпа муравьев, которые вползают на дерево, а через час возвращаются с наполненными зобиками. Camponotus vividus устраивает свои расположенные группами гнезда, расширяя ходы, просверленные в ветках жуками, и собирает корм в лесном пологе на площади в сотни квадратных метров, постепенно сменяя ярусы, а иногда фуражируя даже на земле. Он посещает белокрылок, собирая у них падь, а также собирает различные вещества, выделяемые растениями, в особенности сухую белую смолу; фактически 90% сборов этого муравья — материал растительного происхождения. Пики активности приходятся на 6—8 часов и на 16—18 часов; в промежутках активность падает на 20%. Температура никогда не заставляет этих муравьев прекратить фуражировку; они перестают собирать пищу только в случае дождя. Таким образом, у Camponotus в эксосистеме тропической саванны наблюдается несколько различных образов жизни (Levieux, Louis, 1975).
Столь же четкая экологическая дифференциация выявляется у видов термитов, относящихся к одному роду. Sands (1965) изучал пять видов Trinervitermes (Nasutitermitinae) в западноафриканской саванне, где в августе количество осадков достигает максимума (112—122 мм), а в декабре трава выгорает. Sands обследовал три биома: редколесье с травами высотой 1—2 м, кустарник со сходными травами и область с одними только густыми высокими травами. Ввиду обилия злаков, которыми питаются все эти пять видов, конкуренция между ними за пищу маловероятна, но Sands обнаружил, что они встречаются в разных зонах и различаются по устройству гнезд. T. geminatus живет на открытых солнечных участках в агрегированных гнездовых холмиках и хорошо приспособлен к своему местообитанию и климату. T. trinervius не занимается активной фуражировкой и лучше чувствует себя в более влажной саванне с двумя пиками осадков, к которым лучше адаптирован его жизненный цикл. T. togoensis явно неспособен строить для себя прочные холмики и селится в старых холмиках T. geminatus и Cubitermes.
T. oeconomus и T. occidentals имеют светлую окраску и чувствительны к солнечному свету; они непрерывно собирают злаки, тогда как другие виды создают запасы, которые изолируют гнездо от перегрева на солнце и даже от огня. Больше половины гнезд T. oeconomus находятся в холмиках Macrotermes. Sands приходит к выводу, что различия в гнездостроительных способностях, в способах фуражировки и кормления (хотя не обязательно в пище) и адаптация к климату дают возможность этим видам сосуществовать в грассленде, где очень мало деревьев. Различия в поведении при выборе местообитания обеспечивают пространственную разобщенность видовых популяций, и эти термиты проявляют агрессивность лишь при случайных встречах с другим видом.
Microtermes (Macrotermitinae) — мелкие термиты, которые живут под землей и выращивают грибы; они строят камеры диаметром 3—4 см, соединенные подземными галереями. Wood (1981) в Мокве (![]() Нигерия, 9° с.ш.) изучал семь видов этого рода, которые репродуктивно изолированы. Брачный лёт у большинства видов происходит после дождя, но в разные сроки (через 1, 2 или 3—4 дня) и в характерное для каждого вида время дня или ночи. У некоторых видов половые особи находят друг друга, собираясь у заметных деревьев, а у других самки привлекают самцов, сидя на растениях.
Нигерия, 9° с.ш.) изучал семь видов этого рода, которые репродуктивно изолированы. Брачный лёт у большинства видов происходит после дождя, но в разные сроки (через 1, 2 или 3—4 дня) и в характерное для каждого вида время дня или ночи. У некоторых видов половые особи находят друг друга, собираясь у заметных деревьев, а у других самки привлекают самцов, сидя на растениях.
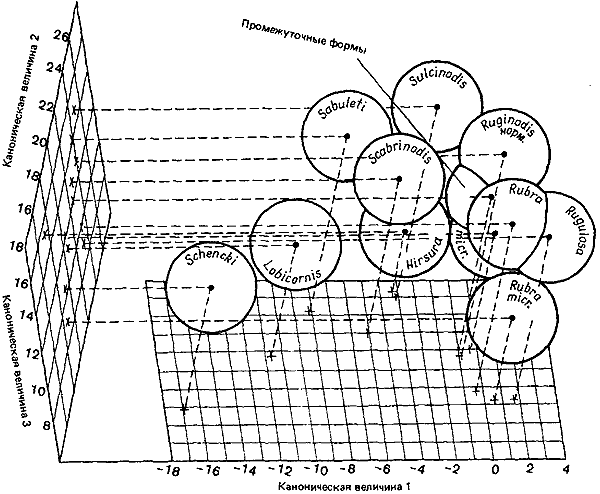
Рис. 88. Биометрический анализ 10 европейских видов Myrmica, включая микрогин (micr.)
живут колониями (пчелы, муравьи, термиты, некоторые осы). В состав колонии, напр. у термитов, входят половые особи (самцы и матки обеспечивают размножение), солдаты (защиту) и рабочие (собирание корма, строительство гнезда и т. д.). Солдаты и рабочие бесплодны; половые особи и солдаты сами не способны питаться; таким образом, члены колонии полностью зависят друг от друга. Разделение функций и сложные взаимодействия разных особей в колониях общественных насекомых обеспечиваются рядом особенностей в их строении и поведении; последнее регулируется с помощью выделяемых ими химических веществ (феромонов).
Всего 134 вида.
совокупность особей одного вида, длительно занимающая определенное пространство и воспроизводящая себя в течение большого числа поколений. В современной биологии популяция рассматривается как элементарная единица процесса эволюции, способная реагировать на изменения среды перестройкой своего генофонда.
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
наличие в семье нескольких яйцекладущих самок. Смотри: моногиния, олигогиния.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
наличие в семье одной яйцекладущей самки. Смотри: полигиния, олигогиния.
Subspecies: Myrmica scabrinodis ussuriensis, Kuznetsov-Ugamsky
сладкая густая жидкость, выделяемая тлями, червецами и другими насекомыми, питающимися соком растений. Медоносная пчела собирает падь с листьев и перерабатывает в падевый мед.
(от греч. oikos дом, жилище, местопребывание и ...логия), наука об отношениях живых организмов и образуемых ими сообществ между собой и с окружающей средой. Термин «экология» предложен в 1866 Э. Геккелем. Объектами экологии могут быть популяции организмов, виды, сообщества, экосистемы и биосфера в целом.
 Myrmica rubra (Linnaeus, 1758) — рыжая мирмика
Myrmica rubra (Linnaeus, 1758) — рыжая мирмика Lasius flavus (Fabricius, 1782) — желтый луговой (садовый, земляной) муравей
Lasius flavus (Fabricius, 1782) — желтый луговой (садовый, земляной) муравейВсего 106 видов.
 Messor capitatus (Latreille, 1798)
Messor capitatus (Latreille, 1798) Messor structor (Latreille, 1798) — степной муравей-жнец
Messor structor (Latreille, 1798) — степной муравей-жнецСтарое название: Messor rufitarsis darianus Pisarski
Старое название: Pheidole arenarum Ruzsky
организационно оформленная мультифункциональная часть семьи (подсистема), имеющая постоянный состав и способная перейти к самостоятельному существованию.
форма поликалии, при которой семья рассредоточена по готовым камерам и небольшим гнездам, не имеющим постоянного состава. Функциональной дифференциации таких гнезд и камер нет.
Всего 427 видов.
Myrmicinae — самое большое подсемейство муравьев, включающее более 2 тыс. видов, относящихся более чем к 150 родам.
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
сборное название насекомых подотряда жалящих перепончатокрылых (кроме пчел и муравьев). Длина до 40 мм (шершни). Ок. 23 тыс. видов. Распространены широко. Опылители растений. Личинки некоторых ос паразиты других насекомых.
Всего 100 видов.
Одно из крупных подсемейств, насчитывает около 1,5 тыс. видов, относящихся к 44 родам. Широко распространены на всем земном шаре, довольно обычны в тропиках и занимают доминирующее положение в умеренных широтах.
 Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant
Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden antподотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
 Lasius alienus (Foerster, 1850) — бледноногий муравей, бледный садовый муравей, полевой муравей, cornfield ant
Lasius alienus (Foerster, 1850) — бледноногий муравей, бледный садовый муравей, полевой муравей, cornfield antв биологии (от др.-греч. πολύμορφος - многообразный) - способность некоторых организмов существовать в состояниях с различной внутренней структурой или в разных внешних формах.
У муравьев это проявляется в том, что представители разных каст морфологически очень сильно отличаются друг от друга. Отсюда условное деление на «рабочих» и «солдат».
Всего 965 видов.
общая масса особей одного вида, группы видов или сообщества в целом на единицу поверхности или объема местообитания. Биомасса растений называется фитомассой, биомасса животных зоомассой.
участок, используемый муравьями для добычи пищи.
отряд общественных насекомых. Общины, разделенные на касты, состоят из крылатых и бескрылых особей. Строят подземные и наземные (до 15 м высотой) гнезда (термитники). Ок. 2600 видов, главным образом в тропиках; в России 2 вида: один в районе Сочи, второй, вероятно, завезенный из Японии или Китая, во Владивостоке. Многоядны. Разрушают древесину, портят бумагу, кожу, сельскохозяйственные продукты и др. Термиты, обитающие в почве, полезны как почвообразователи.






