






У термитов (за исключением сем. Termitidae) рабочие особи мало чем отличаются от личинок, если не считать того, что их покровы несколько жестче и слегка пигментированы, а сами они более активны и более общительны. Развитие их протекает постепенно и не сопровождается какими-либо изменениями в строении зачатков крыльев или гонад. Линька создает возможность для роста и развития, хотя эти процессы имеют место не всегда, а «стационарная» линька означает, что число стадий развития может варьировать. Тем не менее существует ряд определенных типов развития.
Например, у Mastotermes darwiniensis, самого примитивного из ныне существующих термитов, имеются пять личиночных стадий и одна стадия рабочего; на этой последней стадии особи окрашены темнее и занимаются фуражировкой. Но это не завершающая стадия, так как рабочая особь может перелинять и превратиться в нимфу с явно выраженными зачатками крыльев, а затем, после трех нимфальных стадий, стать крылатой половозрелой формой с функционирующей половой системой (мужской или женской). Таким образом, всего здесь 10 стадий. Даже нимфа не детерминирована к тому, чтобы непременно превратиться в имаго, так как она может встать на путь развития, приводящий к солдату. Для этого она должна сначала перелинять в промежуточную форму, называемую просолдатом; на стадии просолдата происходит перестройка, а затем, после новой линьки, получается взрослый солдат; у Mastotermes это просто более крупная бескрылая особь с массивными челюстями и недоразвитыми гонадами.
Это превращение, подобно превращению в имаго, сопровождается утратой морфогенетических потенций: состояние солдата резко отличается от метастабильной стадии рабочего, который может оставаться рабочим, проходя «стационарные» линьки, или же, пройдя через нимфальные стадии, превратиться в половую особь или в солдата и закончить на этом свой жизненный цикл. В небольших семьях могут формироваться узкоголовые солдаты, возможно в результате преждевременного метаморфоза, стимулируемого царской парой (Watson, 1974). Для представителей семейства Calotermitidae (например, Calotermes, Zootermopsis) характерен такой же однонаправленный морфогенез; личинки старших возрастов выполняют различные работы, ожидая, когда настанет их очередь превратиться в имаго или солдат; при этом они проходят через несколько нимфальных стадий или через одну стадию просолдата (рис:73). В этом семействе нимфы могут в случае необходимости превращаться в половых особей, бескрылых или с недоразвитыми крыльями, но вполне эффективных в смысле размножения. Нимфы могут также превращаться в солдат.
У Rhinotermitidae (например, у Reticulitermes) еще сохраняются две общие для всех особей личиночные стадии. В дальнейшем единый ствол лабильных в отношении касты особей расщепляется на два. Новой особенностью является ветвь, ведущая к развитию почти исключительно стерильных каст, т. е. рабочих и солдат.
Этот путь развития состоит из пяти личиночных стадий; личинки выполняют различные работы, однако они могут превращаться в просолдат или бескрылых замещающих половых особей, но никогда не превращаются в нимф или имаго. Они явно утратили способность к развитию крыльев, но развитие гонад у них еще возможно. Другая, лабильная, ветвь состоит из шести нимфальных стадий, которые могут превращаться в замещающих половых особей или же полностью завершать цикл — до крылатых имаго; это не рабочие особи, и они не могут развиться в солдат. Однако эта ветвь лабильна в том смысле, что нимфы способны регрессировать в личинок, т. е. утрачивать свои зачаточные крылья; после этого они могут участвовать в работах, а в дальнейшем превращаться в солдат, пройдя переходную стадию просолдата. У большинства Rhinotermitidae переход к развитию по стерильной ветви зависит от времени года и от питания, а также от размеров семьи, но у представителей рода Schedorhinotermes эта ветвь для самцов закрыта, так что все работы в семье и ее защита лежат на самках.
У Termitidae бескрылых или почти бескрылых замещающих половых особей нет — они, видимо, исчезли в процессе эволюции. Очевидно, крылатые имаго могут обеспечить достаточное замещение, несмотря на то, что они появляются только в определенное время года, как у муравьев. Рабочими особями здесь также служат не личинки с затвердевшими покровами, а особи, развивающиеся из личиночных стадий в результате одной — единственной линьки — обычно окончательной, хотя в отличие от имагинальных стадий у этих рабочих сохраняются проторакальные железы. Несмотря на то что гонады у них недоразвиты, особей разного пола можно различить на личиночной стадии, и теперь можно считать установленным, что вероятность развития по стерильному пути каким-то образом связана с полом. После того как этот путь выбран, вероятность превращения в солдат тоже зависит от пола; например, у Nasutitermitinae солдатами обычно становятся самцы, а у Macrotermitinae и Termitinae — самки. У тех видов, у которых представители данной касты бывают двух разных размеров, это чаще обусловлено различием в стадии созревания, чем половыми различиями (Brian, 1979а; Noirot, 1969, 1974).
У Termitidae репродуктивная и стерильная ветви расходятся после всего лишь одной стадии, возможно даже на стадии яйца (Noirot, 1969). На пути развития половых особей имагинальной стадии предшествуют семь стадий (рис:74). У такого примитивного вида, как Amitermes evuncifer, между яйцом и первой личинкой, выполняющей функции рабочего, имеются две личиночные стадии, а затем три или четыре возраста рабочих. Солдаты, развивающиеся из личинок того и другого пола, образуются большей частью из рабочих первого возраста. У Microtermes (Amitermitinae) рабочие мужского пола после двух личиночных стадий проходят еще три стадии, а рабочие женского пола — четыре стадии и в результате бывают крупнее рабочих-самцов; просолдаты тоже дивергируют рано, от рабочих первого возраста. Другие примеры можно найти у Noirot (1974) и Brian (1979а). В общем можно сказать, что у Termitidae дивергенция репродуктивной и стерильной линий происходит рано, возможно на стадии яйца; что личиночных стадий у них меньше, а нимфальных больше; что солдаты развиваются обычно из личинок определенного пола, а рабочие — одинаково из мужских и женских личинок; что особи разного пола и находящиеся на разных стадиях созревания различаются по размерам.
Люшер (Luscher, 1976) обнаружил у Termitidae (Macrotermes subhyalinus) материнский эффект, обусловленный различным содержанием ювенильного гормона (ЮГ) в яйце. В тот период, когда плодущая царица откладывает яйца, дающие нимф, уровень ЮГ в ее крови низок. В отличие от этого у молодых цариц уровень ЮГ в крови высокий, и все отложенные ими яйца развиваются по стерильной линии. Люшер разработал на основе этих данных общую теорию о роли ЮГ в регуляции каст у термитов; согласно этой теории, ЮГ-подобное вещество, выделяемое в пищевой поток парой половых особей, может приостановить образование замещающих особей во время чувствительной стадии цикла линек. Их способность синтезировать ЮГ, несомненно, очень велика; им нужно большое количество этого гормона, чтобы производить яйца, но остается ли его достаточно для циркуляции в семье (рис:75)? Солдаты, по мнению Люшера, выделяют какое-то анти-ЮГ-вещество, прекращающее дальнейшее образование солдат; убедительно показано, что добавление аналогов ЮГ к культурам термитов, содержащим личинок в чувствительной стадии (первые 6 дней третьей личиночной стадии у M. michaelseni) (согласно Okot-Kotber, 1980), повышает формирование просолдат (Brian, 1979а; Nijhout, Wheeler, 1982). Можно думать, что ЮГ делает возможным дальнейший аллометрический рост, ведущий к характерной гипертрофии головы. Высказано предположение (Nijhout, Wheeler, 1982), что сроки выбросов ЮГ в кровоток важнее, чем общее его содержание в крови.
Данных о морфогенезе каст у примитивных муравьев пока еще мало. У Odontomachus ![]() (Ponerinae) количество пищи может до очень поздних стадий влиять на кастовую дифференциацию; здесь существует один главный лабильный путь развития, но мы не знаем, не отходит ли от него ветвь на более ранней стадии или даже не бывает ли определенного кастового «уклона», заложенного матерью в яйцо. У Myrmica действуют те же регулирующие факторы, что и у
(Ponerinae) количество пищи может до очень поздних стадий влиять на кастовую дифференциацию; здесь существует один главный лабильный путь развития, но мы не знаем, не отходит ли от него ветвь на более ранней стадии или даже не бывает ли определенного кастового «уклона», заложенного матерью в яйцо. У Myrmica действуют те же регулирующие факторы, что и у![]() Ponerinae, и, кроме того, существует несколько способов, при помощи которых самки могут управлять этим процессом. Одна, а может быть, и две линьки заменены пластификацией кутикулы, и в регулировании морфогенеза каст участвуют железы.
Ponerinae, и, кроме того, существует несколько способов, при помощи которых самки могут управлять этим процессом. Одна, а может быть, и две линьки заменены пластификацией кутикулы, и в регулировании морфогенеза каст участвуют железы.
Можно выделить два дискретных и взаимоисключающих физиологических состояния, различающихся по соотношению скоростей роста «зародыша имаго и роста личинки в целом (под «зародышем» здесь имеется в виду совокупность имагинальных дисков, имеющихся в личинке). При формировании репродуктивной самки, соответствующем обычному процессу развития насекомого, «зародыш» растет медленно и большая часть пищи, поглощаемой личинкой, откладывается в жировом теле. Это продолжается до тех пор, пока головной мозг личинки не переместится в грудной отдел, где вокруг него формируется голова взрослой особи. После этого «зародыш» растет изометрически, у него развиваются крылья, разделенные яичники, сегментированные усики и ноги; начинается видимая дифференцировка всех частей организма, необходимых взрослой репродуктивной самке.
Эти процессы становятся возможными в период прохладной погоды (обычно зимой), когда возобновляется активность corpora allata. Развитие «зародыша» метастабильно и может быть нарушено голоданием при температуре выше 23°C. «Зародыш» увеличивается в размерах и изменяет форму: увеличиваются почки ног, но не почки крыльев и не зачатки яичников, что нетрудно увидеть сквозь кутикулу личинки. В целом это приводит к увеличению относительной массы тканей «зародыша» в личинке. Происходящие изменения необратимы и сопровождаются утратой морфогенетических потенций: детерминируется развитие рабочей особи. Быстрый переход «зародыша» из одной формы в другую — это переход между двумя стабильными состояниями, и промежуточных форм в естественных условиях не образуется, если не считать тех случаев, когда под влиянием паразитов нарушается питание.
В процессе роста «зародыша» рабочей особи крылья остаются небольшими, а во время метаморфоза вообще исчезают, включаясь в наружный скелет; маленький гермарий не распадается на трубочки, как у репродуктивной самки, а соединяется с яйцеводом, образуя яичник с одной яйцевой трубочкой (рис:76).
А чем объясняется меньшая величина рабочих особей? Освободившись от крыльев и яичников и увеличившись в размерах, «зародыш» продолжает свое развитие, и питание личинки прекращается на 2—3 дня раньше, чем это бывает при развитии репродуктивной самки; в результате рабочие особи оказываются мельче. Детерминация развития в направлении рабочей особи может быть сравнительно поздним отклонением от лабильной линии, ведущей к развитию репродуктивной самки (рис:77); оно может произойти уже после того, как личинка накопила питательные вещества, диапаузировала, перезимовала и вновь обрела способность к росту; таким образом, здесь проявляется известное сходство с нимфой Schedorhinotermes, которая регрессирует в рабочую особь, хотя, конечно, при этом не бывает линьки и все описанные изменения происходят внутри личиночной кутикулы. Между тем многие рабочие особи (возможно, большинство их) образуются из «зародышей», дивергировавших от лабильного ствола задолго до фазы накопления запасов перед диапаузой; в их число входят все рабочие особи, происходящие из яиц, детерминированных к развитию рабочих.
После такой детерминации дифференцировка обычно замедляется, личинки перестают кормиться и внезапно метаморфизируют; дифференцировка «зародыша» и его превращение в куколку внутри личиночных покровов после прекращения питания — вполне определенный процесс, и он может занимать при развитии рабочей особи столько же времени, что и при развитии репродуктивной самки. Тем не менее детерминацию к развитию рабочей особи можно рассматривать как шаг к метаморфозу, поскольку она вызывает относительный рост «зародыша» внутри личинки, сокращает время роста и тем самым уменьшает окончательные размеры рабочей особи. Это безусловно преждевременный, но незавершенный метаморфоз, необратимо искажающий развитие «зародыша» (Brian, 1974а); его формирование, по-видимому, запускается каким-то сигналом, исходящим от головного мозга, который подавляет активность corpora allata, не включая функцию проторакальных желез.
У Myrmica, как и у всех муравьев, только самки делятся на два типа, различающиеся по размерам, морфологии, поведению и долговечности. У самцов такого деления нет, и мужские «зародыши» не подвержены такому нарушению развития, как женские; зачатки всех частей имаго морфогенетически связаны между собой, и рост «зародыша» протекает изометрически. Роль ювенильного гормона у обоих полов состоит в том, чтобы ограничивать рост «зародыша» и способствовать запасанию питательных веществ в жировом теле личинки. Перевязка corpora allata у женской личинки, лабильной в отношении касты, ведет к детерминации развития рабочей особи, и ее можно приостановить только с помощью аналогов ювенильного гормона (Brian, 1974b).
У Plagiolepis pygmaea активность corpora allata зависит не от числа, а от величины их клеток (Suzzoni, Grimal, 1980). Итак, у муравьев рабочие особи образуются из «зародыша», который заключен в червеобразной личинке, но созревает и отделяется от нее, еще не достигнув полного развития. Этот процесс обособления от личинки прекращает ее дальнейшее развитие, точно так же как личинки высших термитов навсегда становятся рабочими. В этом смысле рабочие особи муравьев и термитов гомологичны. Различия между ними обусловлены лишь тем, что у термитов ювенильные особи соприкасаются с окружающей средой с самого начала, тогда как у муравьев они вначале заключены во временный «футляр», совершенно отличный от них, освобождение из которого кладет конец пластичности их формы.
Процесс формирования солдат у муравьев все еще остается неясным. У Pheidole линия репродуктивных самок отделена от бесполой линии уже на стадии яйца, на развитие которого влияет (см. выше) концентрация ювенильного гормона в гемолимфе матери (Passera, Suzzoni, 1979; Wheeler, Nijhout, 1981). Личинки, дающие самок, толстые и лишены волосков, а личинки бесполых особей вытянутые и покрыты волосками. Отклонение развития в сторону солдат происходит на последней (третьей) личиночной стадии, на которой кишечник имеет золотистую окраску; личинок солдат можно отличить по их более крупным размерам, а также по тому, что у них имеются почки передней пары крыльев, но нет видимых почек задней пары; у личинок рабочих особей совсем нет крыловых почек (или, в редких случаях, они очень малы). У всех этих каст крыловые почки невелики по сравнению с почками ног, но у куколок, дающих солдат, на месте переднего крыла можно видеть палочковидный выступ (как у «крупных рабочих» и интеркастов Myrmica, рис:78).
Размеры солдат и рабочих не перекрываются. Путем местного введения ювенильного гормона во время чувствительной фазы на последней личиночной стадии можно получить гигантского солдата; такое воздействие приостанавливает начало метаморфоза и создает возможности для дополнительного роста, который носит аллометрический характер и приводит к характерной морфологии солдат. Оно приводит также к увеличению размеров рабочих особей, но лишь незначительному, и создает ситуацию, подобную естественному различию между двумя видами Solenopsis — S. invicta (эквивалент необработанных Pheidole) и S. geminata (эквивалент Pheidole после обработки ювенильным гормоном).
В естественных условиях солдаты формируются только в присутствии самки, при обилии пищи, высокой температуре (26°C и выше) и большом числе рабочих особей. Наличие взрослых солдат подавляет дальнейшее их образование, хотя этот эффект к концу сезона размножения уменьшается. Дело не в том, что солдаты — плохие кормилицы и не могут снабжать необходимой пищей личинок, дающих солдат; наблюдаемый эффект зависит от частоты контактов между особями и, по-видимому, связан со способностью разрушать ювенильный гормон (Edwards, 1982; Wheeler, 1982), как и у термитов. Когда удастся разработать схемы онтогенеза промежуточных типов, интересно будет выяснить, каким образом произошел переход от онтогенеза, свойственного Myrmica, с общим лабильным стволом и двумя главными путями развития (дающими мелких и крупных рабочих), к такой картине, как у Pheidole, у которого пути развития репродуктивных самок и рабочих разделены со стадии яйца, а от линии рабочих ответвляется еще линия сверхкрупных солдат (рис:79) (Brian, 1979а).
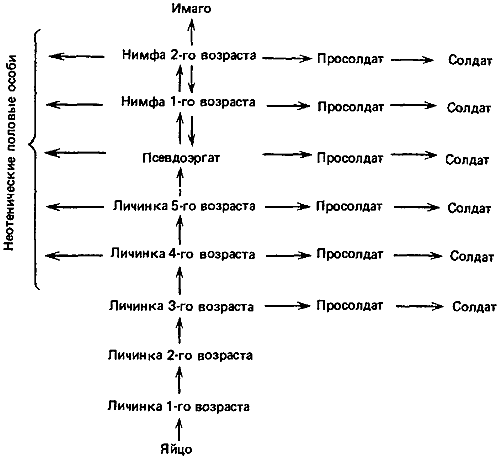
Рис. 73. Возможные пути онтогенетического развития у Calotermes flavicollis. Каждая стрелка соответствует одной линьке; пунктирная стрелка означает варьирующее число линек. (Liischer, 1976.)
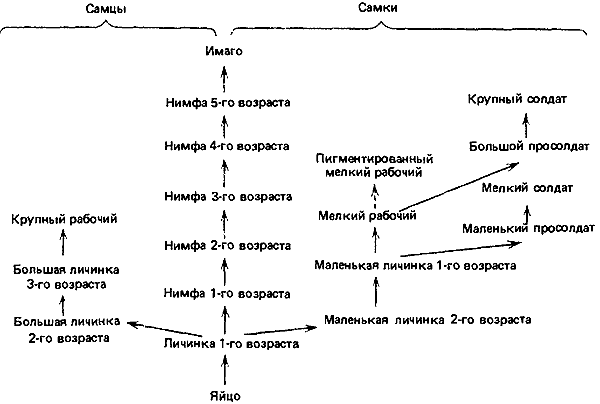
Рис. 74. Возможные пути онтогенетического развития у Macrotermes bellicosus. Каждая стрелка соответствует одной линьке, за исключением пунктирной, которая означает пигментацию без линьки. (Liischer, 1976.)
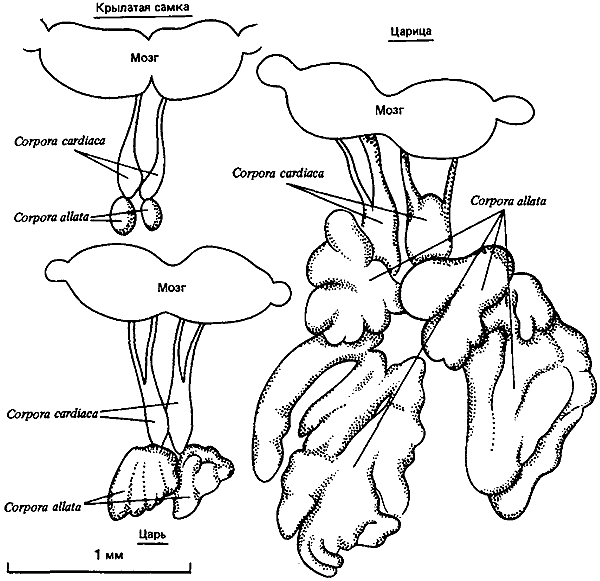
Рис. 75. Эндокринные системы взрослых особей Macrotermes subhyalims. (Luscher, 1976.)
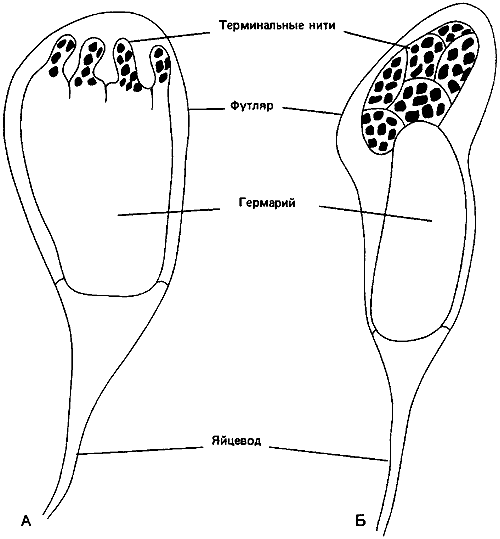
Рис. 76. Яичники личинок Myrmica последнего возраста, развивающихся в репродуктивных самок (А) и в рабочих особей (Б).
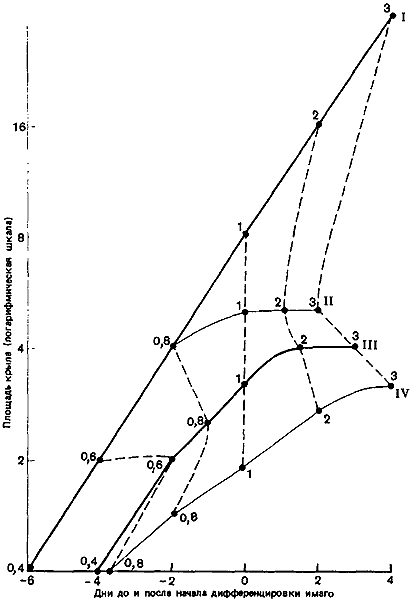
Рис. 77. Рост крыловых имагинальных дисков у личинок Myrmica. I — репродуктивные самки; II — крупные рабочие особи; III — средние рабочие особи; IV — мелкие рабочие особи. Стадии дифференцировки взрослых особей обозначены цифрами 0,6; 0,8; 1; 2; 3. (Brian, 1974а.)
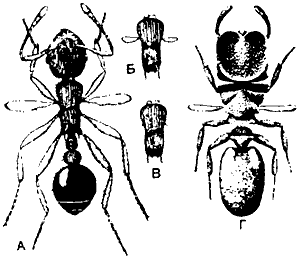
Рис. 78. А—В. Интеркасты Myrmica с палочковидными крыловыми выступами; эти особи развиваются из лабильных личинок, которые не смогли дорасти до настоящих репродуктивных самок и близки к касте, известной как солдаты (например, у Pheidole). Г. Солдат Zacryptoceros со сходными палочковидными крыльями. (Wheeler, 1910.)
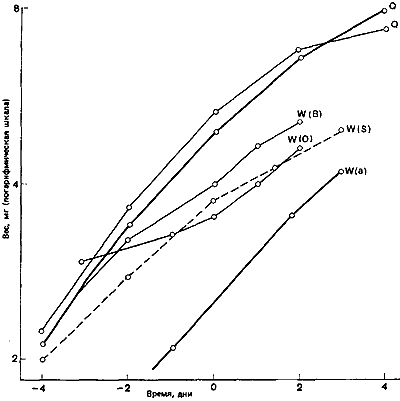
Рис. 79. Зависимость веса растущих личинок Myrmica от времени после начала сегментации ног. Q — самки; W(a) — нормальные рабочие особи средних размеров; W(O) и W(B) — лабильные личинки, которых выращивали в культуре перед началом зимы, когда ростовой потенциал низок; W(S) — лабильные личинки, которых выращивали в культуре после зимнего восстановления ростового потенциала, но при этом сразу после периода голодания при высокой температуре (25°С). W(O, B и S) — прототипы «солдат» (т. е. крупные рабочие особи). (Brian, 1974а.)
отряд общественных насекомых. Общины, разделенные на касты, состоят из крылатых и бескрылых особей. Строят подземные и наземные (до 15 м высотой) гнезда (термитники). Ок. 2600 видов, главным образом в тропиках; в России 2 вида: один в районе Сочи, второй, вероятно, завезенный из Японии или Китая, во Владивостоке. Многоядны. Разрушают древесину, портят бумагу, кожу, сельскохозяйственные продукты и др. Термиты, обитающие в почве, полезны как почвообразователи.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
взрослая особь
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
у муравьев имеется три основные касты: самцы, самки и рабочие (бесплодные модифицированные самки).
необратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.
Довольно примитивная группа муравьев по своему строению и поведению. Рабочие мономорфны, самки по размерам близки к рабочим.
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
Всего насчитывает 549 видов.
«кровь муравьев», прозрачная жидкость кровеносной системы.
Всего 134 вида.
(rp. ontos - сущее, существо) - индивидуальное развитие организма с момента зарождения до смерти.






