






Надсемейство Formicoidea, представленное в современной фауне только семейством муравьев, возникло на рубеже верхнего и нижнего мела около 100 млн. лет назад. Судя по некоторым морфологическим особенностям первых Formicoidea, эти насекомые, внешне похожие на муравьев, еще не были настоящими социальными насекомыми. Древнейшими из известных настоящих муравьев являются еще не описанные самец из янтаря Байкуры (Таймыр, поздний мел) и несколько рабочих особей из палеоцена (сахалинский янтарь). Муравей из Байкуры относится к особому подсемейству, видимо, близкому к Nothomyrmeciinae, а среди десятка особей, найденных в Сахалинском янтаре, одна принадлежит подсемейству![]() Ponerinae (Ectatommini), а остальные —
Ponerinae (Ectatommini), а остальные — ![]() Dolichoderinae. Один из видов, по-видимому относящийся к современному роду Dolichoderus, — типичный дендробионт.
Dolichoderinae. Один из видов, по-видимому относящийся к современному роду Dolichoderus, — типичный дендробионт.
Наиболее изученной среди ископаемых фаун муравьев является фауна Балтийского янтаря (эоцен) (Wheeler, 1914). Здесь найдены почти все современные подсемейства муравьев, однако резко доминируют представители ![]() Dolichoderinae (64,3%) и
Dolichoderinae (64,3%) и![]() Formicinae (32,8% инклюзов). Найдены и многие современные роды, такие, как Formica, Lasius, Camponotus, Iridomyrmex, Aphaenogaster, Cataglyphis, Tapinoma, Prenolepis и др. Показаны четкие связи этой фауны с современными фаунами Па леарктики, Индо-Малайской области и Австралии (Wheeler, 1908b, 1914; Emery, 1920). Хотя образование янтаря шло, видимо, на различных территориях, отличающихся по климату, большое число весьма специализированных дендробионтов (Gesomyrmex, Hypoclinea, Oecophylla, Tetraponera, некоторые Camponotus) указывает на наличие тропических или субтропических лесов. Сведения о фауне муравьев засушливых областей эоцена отсутствуют.
Formicinae (32,8% инклюзов). Найдены и многие современные роды, такие, как Formica, Lasius, Camponotus, Iridomyrmex, Aphaenogaster, Cataglyphis, Tapinoma, Prenolepis и др. Показаны четкие связи этой фауны с современными фаунами Па леарктики, Индо-Малайской области и Австралии (Wheeler, 1908b, 1914; Emery, 1920). Хотя образование янтаря шло, видимо, на различных территориях, отличающихся по климату, большое число весьма специализированных дендробионтов (Gesomyrmex, Hypoclinea, Oecophylla, Tetraponera, некоторые Camponotus) указывает на наличие тропических или субтропических лесов. Сведения о фауне муравьев засушливых областей эоцена отсутствуют.
Тропическая или субтропическая лесная фауна муравьев олигоцена хорошо описана из отложений Флорисанта (Калифорния, США) (Carpenter, 1930). Здесь, как и в балтийском янтаре, доминируют ![]() Dolichoderinae (63,3%) отпечатков), а среди них — дендробионты Protazteca и
Dolichoderinae (63,3%) отпечатков), а среди них — дендробионты Protazteca и![]() Formicinae (32,8%), но удельный вес
Formicinae (32,8%), но удельный вес![]() Myrmicinae (3,7%) все же выше, чем в балтийском янтаре (1,8%). С фауной Флорисанта сходна еще не описанная фауна позднеолигоценовых отложений Приморского края СССР.
Myrmicinae (3,7%) все же выше, чем в балтийском янтаре (1,8%). С фауной Флорисанта сходна еще не описанная фауна позднеолигоценовых отложений Приморского края СССР.
Муравьи миоцена известны нам главным образом по находкам в янтарях Сицилии (Emery, 1891), сланцах Энигена, Радобоя (Heer, 1849), Ставропольского края и Киргизии (Длусский, 1981). Эти данные позволяют представить, как выглядели тогда фауны разных природных зон на территории нынешней Палеарктики. В янтаре Сицилии найдена фауна тропического или субтропического леса. Из 39 инклюзов 8 (21%) принадлежат специализированным дендробионтам (Cataulacus, Gesomyrmex, Oecophylla, Crematogaster). Почти треть (11 инклюзов) всех найденных муравьев составляют рабочие Anoplolepis labialis /Эмери (Emery, 1891) описал этот вид в роде Plagiolepis, но, судя по его рисунку и описанию, его, несомненно, следует отнести к роду Anoplolepis./ — мелкого (1,5—2 мм) стройного муравья с удлиненными конечностями. Соотношение подсемейств близко к современному соотношению в тропических лесах Азии:![]() Ponerinae — 5%,
Ponerinae — 5%,![]() Myrmicinae — 51%, Dolichoderina — 8%,
Myrmicinae — 51%, Dolichoderina — 8%,![]() Formicinae — 36% инклюзов.
Formicinae — 36% инклюзов.
Структура комплексов муравьев миоценовых отложений Ставропольского края (Вишневая балка; караганский горизонт) весьма сходна со структурой комплексов современных южных лесов умеренного пояса. Здесь доминируют стратобионты Paraphaenogaster, герпетобионты Lasius и дендробионты Camponotus. Видом-доминантом был, очевидно, Liometopum, а геобионтами — два найденных вида![]() Ponerinae. Соотношение семейств близко к современному в лесах Южной Европы: Ponerina — 2,7,
Ponerinae. Соотношение семейств близко к современному в лесах Южной Европы: Ponerina — 2,7,![]() Myrmicinae — 40,
Myrmicinae — 40, ![]() Dolichoderinae — 4,
Dolichoderinae — 4,![]() Formicinae — 53,3% отпечатков (Длусский, 1981). Если судить по рисункам отпечатков (Heer, 1849), фауна Западной Европы в миоцене была в общем сходной с фауной Ставрополья, однако точно этого утверждать нельзя до проведения полной ревизии материалов.
Formicinae — 53,3% отпечатков (Длусский, 1981). Если судить по рисункам отпечатков (Heer, 1849), фауна Западной Европы в миоцене была в общем сходной с фауной Ставрополья, однако точно этого утверждать нельзя до проведения полной ревизии материалов.
В свете вопросов, обсуждаемых в данной работе, наибольший интерес представляют муравьи, найденные в Чон-Тузе (Кочкорский р-н Киргизии; миоцен), поскольку они обитали в районе с засушливым климатом на территории современной Средней Азии. Всего найдено 22 отпечатка, обычно плохой сохранности, по которым оказалось возможным описать лишь три вида: «Ponerites» inversa, Rhytidoponera kirghizorum ![]() (Ponerinae), Kotshkorkia latipes (
(Ponerinae), Kotshkorkia latipes (![]() Dolichoderinae) (Длусский, 1981). Rh. kirghizorum относится к мелким представителям рода (подрод Cholcoponera), многочисленным в Австралии и являющимися стратобионтами. Kotschkorkia — это довольно крупные (около 10 мм) муравьи с плотными покровами, хорошо развитыми глазами и нормальными ходильными конечностями. Они могли быть герпетобионтами или неспециализированными дендробионтами. Относительно P. inversus можно сказать лишь, что это небольшая (6,5 мм) понерина, с ногами и усиками умеренной длины. Из остальных 19 отпечатков 6 принадлежат видам рода Camponotus (пять самцов, длина тела 9—10 мм, и одна самка — 11,5 мм) и 13 — небольшим плохо сохранившимся крылатым муравьям с одночлениковым стебельком и округленным брюшком без перетяжки (т. е.
Dolichoderinae) (Длусский, 1981). Rh. kirghizorum относится к мелким представителям рода (подрод Cholcoponera), многочисленным в Австралии и являющимися стратобионтами. Kotschkorkia — это довольно крупные (около 10 мм) муравьи с плотными покровами, хорошо развитыми глазами и нормальными ходильными конечностями. Они могли быть герпетобионтами или неспециализированными дендробионтами. Относительно P. inversus можно сказать лишь, что это небольшая (6,5 мм) понерина, с ногами и усиками умеренной длины. Из остальных 19 отпечатков 6 принадлежат видам рода Camponotus (пять самцов, длина тела 9—10 мм, и одна самка — 11,5 мм) и 13 — небольшим плохо сохранившимся крылатым муравьям с одночлениковым стебельком и округленным брюшком без перетяжки (т. е. ![]() Dolichoderinae или
Dolichoderinae или![]() Formicinae). Один из этих отпечатков (6мм) явно отличается от остальных; оставшиеся по размерам разбиваются на две неравные группы: восемь отпечатков 5 мм длиной и четыре отпечатка 7—7,5 мм длиной. Это могут быть самцы и самки какого-то массового вида (или рода) типа Lasius или Iridomyrmex.
Formicinae). Один из этих отпечатков (6мм) явно отличается от остальных; оставшиеся по размерам разбиваются на две неравные группы: восемь отпечатков 5 мм длиной и четыре отпечатка 7—7,5 мм длиной. Это могут быть самцы и самки какого-то массового вида (или рода) типа Lasius или Iridomyrmex.
Хотя имеющиеся данные очень скудны, все же на их основании можно утверждать, что структура комплекса муравьев в Чон-Тузе отличалась от комплексов современных пустынь Средней Азии. Во-первых, здесь отсутствовали или были редкими карпофаги, всегда представленные видами![]() Myrmicinae. Во-вторых, нередкими были гео- и стратобионты, совершенно отсутствующие в настоящих пустынях. В то же время здесь не найдены и специализированные дендробионты, характерные для тропических лесов. Самки и рабочие дендробионтов, выгрызающих ходы в древесине, имеют твердый жевательный край мандибул, как правило, хорошо сохраняющийся на отпечатках и часто укороченные конечности, а специализированные виды, живущие в кронах, имеют вытянутое тело и голову и удлиненные конечности. Ни те, ни другие формы в Чон-Тузе не обнаружены.
Myrmicinae. Во-вторых, нередкими были гео- и стратобионты, совершенно отсутствующие в настоящих пустынях. В то же время здесь не найдены и специализированные дендробионты, характерные для тропических лесов. Самки и рабочие дендробионтов, выгрызающих ходы в древесине, имеют твердый жевательный край мандибул, как правило, хорошо сохраняющийся на отпечатках и часто укороченные конечности, а специализированные виды, живущие в кронах, имеют вытянутое тело и голову и удлиненные конечности. Ни те, ни другие формы в Чон-Тузе не обнаружены.
Больше всего фауна Чон-Туза похожа на фауну саванны. В качестве примера для сравнения можно взять хорошо изученные саванны Берега Слоновой Кости (Levieux, 1967, 1973). Наиболее многочисленны здесь виды рода Camponotus, среди которых есть как наземные (C. acvapimensis — самый распространенный вид, до 32% всех гнезд), так и древесные формы и небольшие герпетобионты-зоофаги (Acantholepis, Paratrechina, до 17% всех гнезд). Довольно обычны гео- и стратобионты (Amblyopone, Hypoponera, Mesoponera). Правда, здесь обычны и мелкие![]() Myrmicinae, которые могут быть (питание не изучалось) карпофагами, но численность их в несколько раз меньше, чем численность зоонекрофагов. Гнезда
Myrmicinae, которые могут быть (питание не изучалось) карпофагами, но численность их в несколько раз меньше, чем численность зоонекрофагов. Гнезда![]() Myrmicinae, которые потенциально могут быть карпофагами (Tetramorium, Pheidole), составляли на разных участках от 5 до 30% (среднее 12,5%) всех гнезд. Для сравнения можно указать, что в Сахаре гнезда карпофагов составляют от 36 до 59% (среднее — 52%) от общего числа гнезд муравьев (Delye, 1968), а в саксаульниках Каракумов — от 58 до 72% (Длусский, 1975а).
Myrmicinae, которые потенциально могут быть карпофагами (Tetramorium, Pheidole), составляли на разных участках от 5 до 30% (среднее 12,5%) всех гнезд. Для сравнения можно указать, что в Сахаре гнезда карпофагов составляют от 36 до 59% (среднее — 52%) от общего числа гнезд муравьев (Delye, 1968), а в саксаульниках Каракумов — от 58 до 72% (Длусский, 1975а).
Таким образом, хотя муравьи в ископаемом состоянии представлены довольно полно, вплоть до верхнего миоцена мы не находим ни настоящих пустынных комплексов, ни типично пустынных форм. В какой-то степени это связано с условиями захоронения: ископаемые смолы образовывались в древних лесах тропического, субтропического и, возможно, умеренного пояса, а сланцы — у побережья мелководных озер или морей. В настоящей пустыне остатки насекомых сохраниться не могут. В то же время если представить, что сейчас в Средней Азии идет захоронение насекомых, то даже в пресных озерах, окруженных тугаями, наряду с влаголюбивыми Formica и Tapinoma должны в больших количествах откладываться виды Cataglyphis, Messor и Monomorium.
Отсутствие или большую редкость типично пустынных муравьев в миоценовых отложениях Чон-Туза можно объяснить одной из трех гипотез:
1. Пустынные роды и виды муравьев и пустынные комплексы сформировались в плиоцене (1—10 млн. лет назад).
2. Роды, характерные для пустынных комплексов Средней Азии, возникли на других территориях и в плиоцене или плейстоцене проникли в Среднюю Азию, где из них сформировались пустынные комплексы.
3. Настоящие пустынные виды и пустынные комплексы муравьев на территории Средней Азии сформировались уже к миоцену, но обитали на ограниченных территориях и не были так широко распространены, как в настоящее время.
Все, что мы знаем о темпах эволюции родов муравьев, явно противоречит первой гипотезе. Большинство современных палеарктических родов сформировались уже к эоцену и найдены в балтийском янтаре. Времени, прошедшего с нижнего плейстоцена (1—0,5 млн. лет), достаточно лишь для образования близких видов, отличающихся незначительными признаками (Длусский, 1967а). В то же время типично аридные роды (Messor, Cataglyphis) характеризуются высокой дифференциацией. Имеются и прямые свидетельства против этой гипотезы. В балтийском янтаре найден не слишком специализированный представитель рода Cataglyphis — C. constrictus ( = Camponotus conslrictus Mayr, = Formica constricta Wheeler). Самец миоценового Paraphaenogaster microphthalmus (Длусский, 1981) уже обладал весьма специализированным строением груди, характерным для современных представителей рода Aphaenogaster, хотя и выраженным не столь ярко. На этом основании можно предполагать, что расхождение Messor, самцы которых сохранили нормальную форму груди, и Aphaenogaster произошло раньше середины миоцена.
Основу комплексов в пустынях, как правило, составляют виды родов, распространение которых ограничивается аридными и семиаридными областями (рис:9, рис:14). Если некоторые виды и заходят в гумидные области, то встречаются там только в ксеротермных участках. Все эти роды отличаются от близких к ним родов гумидных областей главным образом комплексом признаков, которые могли выработаться только как адаптация к жизни в аридных условиях. Иногда они сочетаются с признаками, характерными для наиболее архаичных представителей близких родов, обитающих в гумидной зоне.
До недавнего времени считалось (Kusnezov, 1956), что наиболее примитивным родом трибы Formicini, куда относится род Cataglyphis, является Proformica, а в его пределах — подрод Alloformica. Для представителей этого подрода характерно сочетание наиболее архаичных признаков трибы — неспециализированное строение груди и стебелька (также характерно для Polyergus и примитивных Formica), неспециализированные нижнечелюстные щупики (также многие Formica и все Proformica), мандибулы с удлиненным вершинным зубцом (также все Proformica и Cataglyphis), отсутствие вариабельности рабочей маеты и т. д. В 1978 г. в Варганзе мы впервые обнаружили самца Alloformica aberrans, который оказался очень сходным с самцами Cataglyphis и сильно отличался от самцов Proformica и Formica, так что Alloformica следует выделять в самостоятельный род и сближать не с Proformica, а с Cataglyphis. Последние отличаются от Alloformica развитым псаммофором, более стройным телом, удлиненными дыхальцами эпинотума, а у специализированных видов — также и модифицированным стебельком, т. е. признаками, которые выработались как адаптация к аридным условиям. Proformica отличаются от близких к ним примитивных видов Formica, с одной стороны, более архаичным строением мандибул (как и Alloformica и Cataglyphis), а с другой — развитием касты плерэргатов и связанными с ним морфологическими изменениями крупных рабочих (адаптации к семиаридным условиям).
Виды рода Messor, произошедшего от Aphaenogaster, отличаются от видов последнего менее специализированным строением груди самцов, рядом адаптации, связанных с переходом к зерноядности (массивная голова рабочих как результат сильного развития мандибулярных мышц и широкие массивные мандибулы с тупыми зубцами), вариабельностью рабочей касты и более или менее развитым псаммофором (адаптации к аридным условиям). Аналогичные различия мы находим и при сравнении Melophorus с другими родами трибы Melophorini, Myrmecocystus — с другими родами трибы Lasiini или Pogonomyrmex — с другими родами трибы Myrmicini. Отсюда следует, что эти роды могли возникнуть только в аридных или семиаридных областях. Поскольку палеоклиматологические данные свидетельствуют о том, что с мелового периода, т. е. с момента возникновения муравьев, наиболее засушливым климатом отличались приблизительно те же области, что и сейчас, у нас есть все основания полагать, что эти роды возникли в пределах их современных ареалов.
Как показал сравнительно-морфологический анализ рода Messor (Bernard, 1973), ближе всего к исходному прототипу рода (и к роду Aphaenogaster) стоит M. aphaenogasteroides, обитающий в горах Афганистана, и виды группы M. excursionis, ареал которых охватывает пустыни Средней Азии, Афганистана, Казахстана и Центральной Азии, а максимальное число видов отмечено в Каракумах и Кызылкумах. В то же время в Сахаре и Южной Африке обитают наиболее продвинутые представители рода. Аналогичная картина получается и при сравнительно-морфологическом анализе трибы Formicini. Оба известных вида Alloformica сохранились в «полусаваннах» Средней Азии. Ареал наиболее примитивных Cataglyphis (C. emeryi, C. pallida) также ограничен Южным Казахстаном, Средней Азией и Афганистаном. Среди Proformica наиболее примитивная P. splendida, у которой рабочие наименее вариабельны и каста плерэргатов отсутствует, обитает в высокогорных степях Тянь-Шаня.
Таким образом, данные палеонтологии, сравнительной морфологии и зоогеографии свидетельствуют, что формирование типичных для современных пустынь Палеарктики групп муравьев началось гдето на территории, включающей современные Казахстан, Среднюю Азию и Афганистан. Типичные пустынные комплексы должны были, хотя бы в несовершенном виде, сформироваться если не в эоцене, то по крайней мере к миоцену. В то же время в миоцене эти муравьи не были еще слишком распространенными и в Средней Азии доминировали комплексы, характерные для саванн. Логично предположить, что возникновение пустынных комплексов шло в наиболее бедных азональных растительных ассоциациях типа песчаных дюн, солончаков или каменистых склонов со скудной растительностью.
Анализ фаун «полусаванн» Средней Азии, т. е. территорий, наиболее близких по климату к миоценовой саванне, показывает, что только здесь обитает целый ряд эндемичных реликтовых видов. В Бадхызе и Копетдаге встречаются Атblуоропе аппае и Aphaenogaster fabulosa, в «полусаваннах» Таджикистана — Aphaenogaster januschevi и Alloformica nitidior, в верховьях и в долине Кашкадарь — Aphaenogaster raphidiiceps, Alloformica aberrans, Tetramorium striativentre, Leptanilla alexandri и еще не описанные реликтовые виды Ponera и Tetramorium. Особое богатство реликтовыми формами «полусаванн» долины Кашкадарьи, видимо, объясняется изолированностью этой территории. Можно предположить, что некоторые![]() Ponerinae, Aphaenogaster из подрода Dorymyrma, Alloformica и Tetramorium, а также Camponotus из подрода Tanaemyrmex были обычными обитателями (а возможно, и доминирующими группами муравьев) в миоценовой саванне на территории современной Средней Азии и Казахстана.
Ponerinae, Aphaenogaster из подрода Dorymyrma, Alloformica и Tetramorium, а также Camponotus из подрода Tanaemyrmex были обычными обитателями (а возможно, и доминирующими группами муравьев) в миоценовой саванне на территории современной Средней Азии и Казахстана.
Анализируя процессы развития аридных флор, А. Н. Криштофович (1946) выделяет три пути формирования аридной флоры: 1) захват новой аридной области элементами из рядом расположенной области; 2) появление из соседних областей отдельных элементов; 3) автохтонное выявление отдельных элементов. Первый путь является наиболее простым и быстрым. В том случае, если рядом нет областей, откуда могут проникнуть целые растительные формации или их основные элементы, аридные области занимают виды гумидных областей, способные выжить в этих условиях. При этом виды, прежде встречавшиеся редко, могут стать доминантными. Именно так злаки Imperata и Andropogon, прежде обитавшие в лесах, положили начало травянистым формациям. Биоценозы, возникшие таким путем, должны отличаться сравнительной бедностью видового состава, хотя общая численность может быть и большой. Таким образом, на начальном этапе формирования новых растительных формаций (в том числе аридных) происходит перемещение и отбор уже сформировавшихся таксонов, которые дают новые сочетания. Впоследствии на базе этих элементов возникают в процессе эволюции специализированные виды и новые таксоны.
Несомненно, все сказанное относится не только к растительным ассоциациям, но и к любым ассоциациям живых организмов (в том числе и к комплексам муравьев) и к биоценозам вообще. Как формируются первичные комплексы муравьев аридных областей, хорошо видно на примере изолированных азональных ландшафтов, в которых условия сходны с условиями в пустынях, где на основе гумидной или семиаридной фауны формируются обедненные ассоциации муравьев. Одними из таких ландшафтов являются, несомненно, песчаные дюны, где, как и в пустынях, велики максимальные температуры и суточные колебания температуры на почве, растительность скудна, а подстилка и гумусный горизонт отсутствуют.
На дюнах в пойме р. Клязьмы во Владимирской области встречаются всего три вида муравьев: ![]() Lasius niger, Formica cinerea,
Lasius niger, Formica cinerea, ![]() Tetramorium caespitum. Два первых вида являются герпетобионтами-зоонекрофагами, рабочие которых довольно сильно отличаются размерами (свежий вес рабочих 2 и 4,5 мг), причем F. cinerea активна исключительно днем, a
Tetramorium caespitum. Два первых вида являются герпетобионтами-зоонекрофагами, рабочие которых довольно сильно отличаются размерами (свежий вес рабочих 2 и 4,5 мг), причем F. cinerea активна исключительно днем, a ![]() L. niger — как днем, так и ночью. Питание
L. niger — как днем, так и ночью. Питание ![]() T. caespitum довольно разнообразно, но значительную часть рациона составляют семена.
T. caespitum довольно разнообразно, но значительную часть рациона составляют семена.
На песчаных пастбищах в Сев. Японии (Абе, 1971) обитают четыре массовых вида муравьев — Messor aciculatus, ![]() Tetramorium caespitum, Formica japonica и Camponotus japonicus. Первый вид, как и все виды рода, является карпофагом-герпетобионтом. Три остальных питаются различными беспозвоночными, но
Tetramorium caespitum, Formica japonica и Camponotus japonicus. Первый вид, как и все виды рода, является карпофагом-герпетобионтом. Три остальных питаются различными беспозвоночными, но ![]() T. caespitum осенью в основном переходит на питание семенами. Средний вес добычи всех четырех видов статистически достоверно отличается. Автор ничего не пишет о суточной ритмике активности, но по моим наблюдениям в Приморском крае M. aciculatus и F. japonica активны исключительно в светлое время суток, а у C. japonicus летом максимальная активность наблюдается ночью.
T. caespitum осенью в основном переходит на питание семенами. Средний вес добычи всех четырех видов статистически достоверно отличается. Автор ничего не пишет о суточной ритмике активности, но по моим наблюдениям в Приморском крае M. aciculatus и F. japonica активны исключительно в светлое время суток, а у C. japonicus летом максимальная активность наблюдается ночью.
На приморских дюнах на юге Приморского края (окр. г. Хасан) доминирующими видами муравьев были Messor aciculatus и Formica picea, реже встречались F. japonica, ![]() Lasius alienus и C. japonicus. Все эти виды являются герпетобионтами, причем первый — карпофагом, а остальные — зоонекрофагами. Все зоонекрофаги отличаются размерами рабочих особей: средний вес рабочего
Lasius alienus и C. japonicus. Все эти виды являются герпетобионтами, причем первый — карпофагом, а остальные — зоонекрофагами. Все зоонекрофаги отличаются размерами рабочих особей: средний вес рабочего ![]() L. alienus — 1,9 мг, F. picea — 4,5 мг, F. japonica — 8,6 мг и C. japonicus — 25 мг. F. picea и F. japonica активны исключительно днем, но вместе почти не встречаются, так как предпочитают разные стации (F. japonica — с богатой растительностью и более плотной почвой). C. japonicus, как отмечалось выше, активен в основном ночью. Активность
L. alienus — 1,9 мг, F. picea — 4,5 мг, F. japonica — 8,6 мг и C. japonicus — 25 мг. F. picea и F. japonica активны исключительно днем, но вместе почти не встречаются, так как предпочитают разные стации (F. japonica — с богатой растительностью и более плотной почвой). C. japonicus, как отмечалось выше, активен в основном ночью. Активность ![]() L. alienus не изучалась.
L. alienus не изучалась.
В Алайской долине на высоте 2600 м (окр. Дарауткургана) на щебнистом плато с подушкообразными ксерофитами и очень редкой (проективное покрытие 0,3) травянистой растительностью обитали всего три вида муравьев — крупный дневной зоонекрофаг ![]() Cataglyphis aenescens, мелкий дневной зоонекрофаг Proformica coriacea и мелкий карпофаг
Cataglyphis aenescens, мелкий дневной зоонекрофаг Proformica coriacea и мелкий карпофаг ![]() Tetramorium caespitum, причем первые два вида были весьма многочисленными, а третий — редок. На луговостепных участках, которые окружали плато, все три вида также были обнаружены, но доминировали в этих ассоциациях несколько видов Myrmica и Formica lemani; встречались также Formica picea, Leptothorax spp., Lasius flavescens, Camponotus buddhae.
Tetramorium caespitum, причем первые два вида были весьма многочисленными, а третий — редок. На луговостепных участках, которые окружали плато, все три вида также были обнаружены, но доминировали в этих ассоциациях несколько видов Myrmica и Formica lemani; встречались также Formica picea, Leptothorax spp., Lasius flavescens, Camponotus buddhae.
Сходная картина наблюдается и при сравнении типичных степных ассоциаций (Кургальджино) и участков с редкими кустиками полыни и злаков с проективным покрытием 0,4—0,6 (Нагыман) в Кургальджинском заповеднике (табл.3). Из всего разнообразия степных видов на участках со скудной растительностью встречаются только три: крупный дневной герпетобионт-зоонекрофаг ![]() Cataglyphis aenescens, мелкий зоонекрофаг Leptothorax serviculus (в основном в кустиках полыни) и
Cataglyphis aenescens, мелкий зоонекрофаг Leptothorax serviculus (в основном в кустиках полыни) и ![]() Tetramorium caespitum, в питании которого значительную долю составляют семена.
Tetramorium caespitum, в питании которого значительную долю составляют семена.
Все приведенные примеры показывают, что как только возникает ассоциация, напоминающая пустыню, в ней из имеющихся видов формируется комплекс муравьев, структура которого представляет собой упрощенный вариант структуры типичных пустынных комплексов и отличается от структуры комплексов окружающих эту ассоциацию зональных ландшафтов. Чаще всего такой комплекс складывается из двух видов герпетобионтов-зоонекрофагов, рабочие которых отличаются размерами, и одного вида, в питании которого значительную долю составляют семена.
По всей видимости, именно так складывались первичные пустынные Комплексы в эоценовых саваннах на территории современной Средней Азии. Можно предположить, что первыми поселенцами в азональных ландшафтах со скудной растительностью здесь были дневные зоонекрофаги Alloformica (предки Cataglyphis), ночные зоофаги Camponotus и Aphaenogaster, склонные к зерноядности (предки Messor), а возможно, также и мелкие муравьи, склонные к зерноядности, — предки современных Monomorium barbatulum и Tetramorium striativentre.
Это предположение в какой-то степени совпадает с мнением Н. Н. Кузнецова (1926а), считавшего, что древнейшими в фауне Средней Азии являются индийские элементы, в частности пустынные виды Camponotus, представляющие собой остатки существовавшей здесь мезофильной субтропической фауны /В более ранней работе (Кузнецов, 1923) он считал, что эти виды появились в Средней Азии после миоцена./. Необходимо только уточнить, что термин «индийский» не совсем точен. Речь идет не о мигрантах из Индо-Малайской области, а об остатках древней фауны этого района. Многие роды и группы, представленные в настоящее время только в Индо-Малайской области, в эоцене были распространены вплоть до Скандинавии (балтийский янтарь) (Emery, 1920). Несомненно справедливо и мнение Н. П. Кузнецова (1923, 1926а) об автохтонном происхождении среднеазиатских Proformica. В то же время его утверждение о средиземноморском происхождении Messor и Cataglyphis явно ошибочно.
Обсуждая вопрос о происхождении пустынных фаун, К. Эмери (Emery, 1920) и Н. Н. Кузнецов (Kusnezov, 1956) обращали внимание на высокий эндемизм пустынных муравьев и на совпадение современных ареалов родов пустынных муравьев и близких к ним родов, обитающих в гумидных и семиаридных областях, которые скорее всего являются их предками (рис:9, рис:12, рис:14). Действительно, многие черты сходства и различия фаун основных пустынных областей хорошо объясняются различиями в фауне гумидных областей, на базе которых шло формирование пустынных комплексов. Так, например, род Pheidole, представители которого предрасположены к карпофагии, распространен в тропиках и субтропиках всех континентов и его виды выступают как мелкие карпофаги почти во всех пустынях Земли (рис:14). Ареал Aphaenogaster, также предрасположенных к переходу к карпофагии, охватывает Голарктику и Индо-Малайскую область, и соответственно родственные ему роды Veromessor, Novomessor и Messor возникли только в пустынях Неарктики и Палеарктики (рис:14). Род Iridomyrmex, распространенный в Неотропической Австралийской и Индо-Малайской областях, дал два близких рода: один в пустынях Северной Америки (Conomyrma), другой (Dorymyrmex) в пустынях Южной Америки, а в пустынях Австралии — ряд видов, приспособленных к жизни в аридных условиях. Большинство видов рода Manica, ближайших родственников Pogonomyrmex пустынь Северной Америки, также встречается в Северной Америке; лишь один вид этого рода обитает в Европе. Ареал Notoncus, которые скорее всего были предками Melophorus австралийских пустынь, ограничен территорией Австралии.
В то же время, как справедливо отмечал Н. Н. Кузнецов (Kusnezov, 1956), способность переходить к жизни в пустыне не является специфическим свойством каких-либо родов. Так, например, ареал рода Camponotus охватывает все континенты, кроме Антарктиды, причем на севере и юге его границы почти совпадают с границами распространения муравьев. Представители этого рода — характернейший элемент пустынь Палеарктики, Южной Америки, Южной Африки и Австралии. В то же время в Северной Америке пустынные Camponotus отсутствуют, хотя виды этого рода обычны в гумидных и семиаридных ландшафтах континента и известны из олигоценовых отложений Северной Америки. Точно так же Monomorium обитают в гумидной зоне Старого и Нового Света, но в палеарктических пустынях это обычнейшие карпофаги, а в пустынях Нового Света виды этого рода отсутствуют. Lasiophanus — близкие родственники Melophorus австралийских пустынь, обитающие в Южной Америке, встречаются только в районах с влажным климатом.
Сопоставление данных об ассоциациях муравьев азональных песчаных дюн в разных природных зонах показывает, что во всех случаях входящие в них виды обладают рядом общих черт. С одной стороны, это эврибионты с очень пластичным поведением, встречающиеся буквально во всех типах экосистем. К таким видам относятся, например, ![]() Lasius niger в лесной зоне, Lasius alien us и
Lasius niger в лесной зоне, Lasius alien us и ![]() Tetramorium caespitum в степи и лесостепи. Для них характерен довольно высокий уровень социальной организации, с которым связана высокая пластичность гнездостроения и разнообразие способов использования кормового участка. Однако часто доминируют в этих ассоциациях виды, которые не характерны для зональных ландшафтов данной природной зоны и в типе приурочены к нарушенным азональным или интразональным ландшафтам. Характерный пример — Formica picea в Приморском крае. Этот вид встречается здесь, помимо дюн, в тальвегах рек и по морским побережьям. Camponotus japonicus, другой типичный обитатель дюн в Приморье, обычен также на морских побережьях и на каменистых склонах сопок Formica cinerea и
Tetramorium caespitum в степи и лесостепи. Для них характерен довольно высокий уровень социальной организации, с которым связана высокая пластичность гнездостроения и разнообразие способов использования кормового участка. Однако часто доминируют в этих ассоциациях виды, которые не характерны для зональных ландшафтов данной природной зоны и в типе приурочены к нарушенным азональным или интразональным ландшафтам. Характерный пример — Formica picea в Приморском крае. Этот вид встречается здесь, помимо дюн, в тальвегах рек и по морским побережьям. Camponotus japonicus, другой типичный обитатель дюн в Приморье, обычен также на морских побережьях и на каменистых склонах сопок Formica cinerea и ![]() Tetramorium caespitum в лесной зоне Европейской части СССР и Messor aciculatus в Приморье обитают, только в азональных ксеротермных ландшафтах. Как правило, в гумидной зоне обитает несколько видов со сходными спектрами белкового питания, потенциально способных освоить азональные ландшафты со скудной растительностью, имитирующие пустыню, но осваивает их лишь один из них, обычно наиболее термофильный. Так, в лесной зоне европейской части СССР четыре вида Formica потенциально способны существовать на песчаных дюнах — F. cinerea,
Tetramorium caespitum в лесной зоне Европейской части СССР и Messor aciculatus в Приморье обитают, только в азональных ксеротермных ландшафтах. Как правило, в гумидной зоне обитает несколько видов со сходными спектрами белкового питания, потенциально способных освоить азональные ландшафты со скудной растительностью, имитирующие пустыню, но осваивает их лишь один из них, обычно наиболее термофильный. Так, в лесной зоне европейской части СССР четыре вида Formica потенциально способны существовать на песчаных дюнах — F. cinerea, ![]() F. cunicularia,
F. cunicularia, ![]() F. rufibarbis и F. picea. Но живет на дюнах здесь только один из них — F. cinerea. В Казахстане, где практически отсутствуют F. cinerea, в азональных формациях со скудной растительностью часто поселяется
F. rufibarbis и F. picea. Но живет на дюнах здесь только один из них — F. cinerea. В Казахстане, где практически отсутствуют F. cinerea, в азональных формациях со скудной растительностью часто поселяется ![]() F. cunicularia, а в Туве (Жигульская, 1968) и в Приморском крае, где отсутствуют оба эти вида, — F. picea.
F. cunicularia, а в Туве (Жигульская, 1968) и в Приморском крае, где отсутствуют оба эти вида, — F. picea.
По всей видимости, закономерности, которые мы наблюдаем при формировании коадаптивных комплексов муравьев пустынь, — лишь частный случай более общих закономерностей. В настоящее время известно лишь несколько работ, в которых делается попытка реконструировать происхождение не отдельных видов, а целых фаунистических комплексов. М. С. Гиляров (1951, 1954), изучая историю формирования сорной растительности и почвенной фауны полей, показал, что они произошли от флоры и фауны выбросов земли из нор сурков и сусликов целинной степи. На таких выбросах формировалась своеобразная растительность и почвенная фауна, которые легко переносили постоянное перемешивание почвы. Конкурентоспособность этих видов в целинной степи была невелика, но после распашки степей они широко расселились и стали обычными обитателями полей.
Э. О. Вильсон (Wilson, 1961) на основании изучения муравьев Меланезии пришел к выводу, что формирование лесных фаун океанических островов шло по следующей схеме: вначале на базе фауны континента или большого острова (Новая Гвинея) формируется своеобразный комплекс видов прибрежных ассоциаций. Виды, входящие в этот комплекс, обладают высокой пластичностью, которая обеспечивает им возможность существования в нестабильных условиях побережий. Потенциально эти виды легче всего расселяются, и поэтому именно они заселяют океанические острова, лишенные собственной фауны, причем здесь они поселяются не только на побережье, но и в лесах внутри островов. Впоследствии виды претерпевают автохтонную переработку и на разных группах островов формируются эндемичные лесные фауны.
A. С. Северцов (1978), реконструировав происхождение наземных позвоночных, приходит к заключению, что наземной фауне стегоцефалов должна была предшествовать своеобразная фауна обитателей уреза воды — форм, промежуточных между типичными стегоцефалами и кистёперыми рыбами. По его представлениям, именно в этих нестабильных условиях могли сформироваться характерные для амфибий способы дыхания, захватывания пищи и передвижения.
B. В. Жерихин (1978), изучив огромный материал по меловым ископаемым насекомым, показал, что на рубеже верхнего и нижнего мела за сравнительно короткий срок мезозойские группы насекомых почти полностью заменяются кайнозойскими. Причину мелового кризиса автор объясняет тем, что на смену растительности мезофита пришли покрытосеменные растения. Первично эти растения занимали нарушенные экосистемы и не входили в состав нормальных сукцессионных рядов, т. е. имели «сорный» характер. С этими растениями были связаны определенные группы насекомых. Довольно быстро (в геологическом масштабе) покрытосеменные растения вытеснили растительность мезофита, и насекомые, связанные с «сорной» растительностью, широко распространились и начали быстро эволюционировать.
Таким образом, во всех случаях, когда детально анализировалась история формирования фаунистических комплексов /Интересно отметить, что в перечисленных выше работах авторы приходят к сходным выводам, используя разные методы. Э. О. Вильсон пользовался данными таксономии и биогеографии, А. С. Северцов — сравнительной морфологии, В. В. Жерихин — палеонтологии./, оказывалось, что вначале из имеющейся в наличии фауны формировались комплексы видов, связанных с нестабильными азональными экосистемами (прибрежные участки, дюны и т. д.). Затем после изменения условий эти специфические фауны получали преимущества и на их основе возникали сравнительно быстро новые комплексы, слагающиеся уже из специализированных видов. Можно предполагать, что этот путь является если не обязательным, то во всяком случае весьма распространенным.
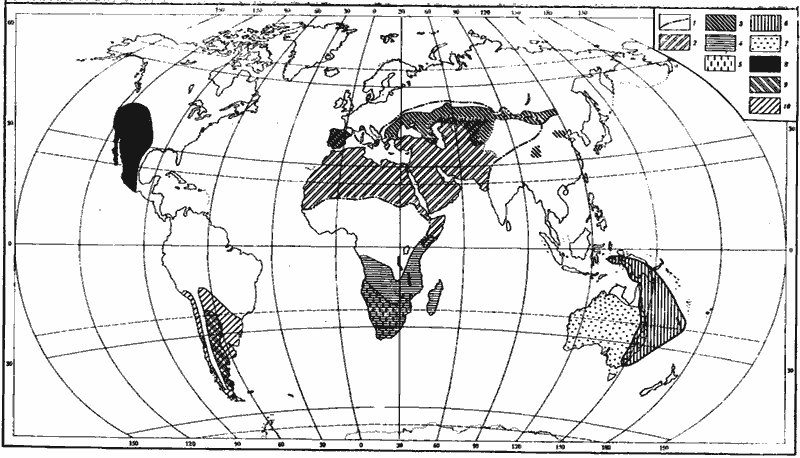
Рис. 9. Географическое распространение основных родов пустынных дневных зоонекрофагов и некоторых «медовых муравьев»
1 — граница распространения Caiaglyphis; 2 — зона, в пределах которой обитает более одного вида Cataglyphis; 3 — Proformica (распространение в КНР, Иране и Турции не изучено; в Афганистане, Индии и Пакистане отсутствуют); 4 — Ocymyrmex; 5 — Anoplolepis (кроме A. longipes, завезенного во многие тропические страны); 6 — Leptomyrmex; 7 — Melophorus; 8 — Myrmecocystus; 9 — Dorymyrmex; 10 — Araucomyrmex
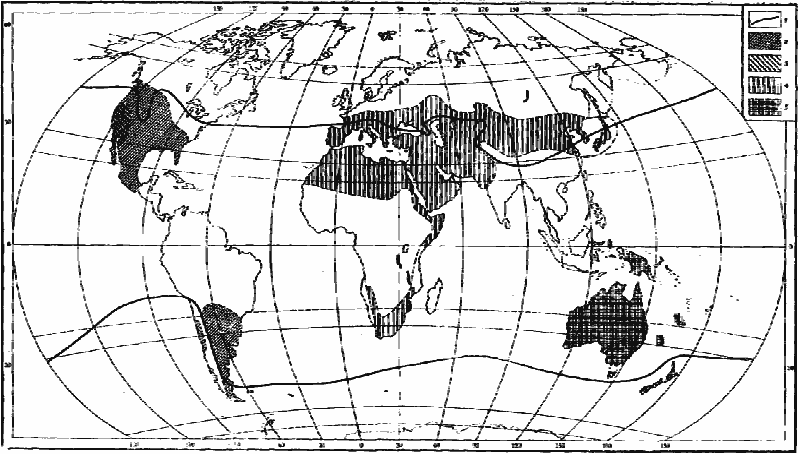
Рис. 14. Распространение некоторых родов
1 — северная и южная границы распространения Pheidole;
2 — Pogonomyrmex;
3 — Veromessor;
4 — Messor;
5 — Chelaner
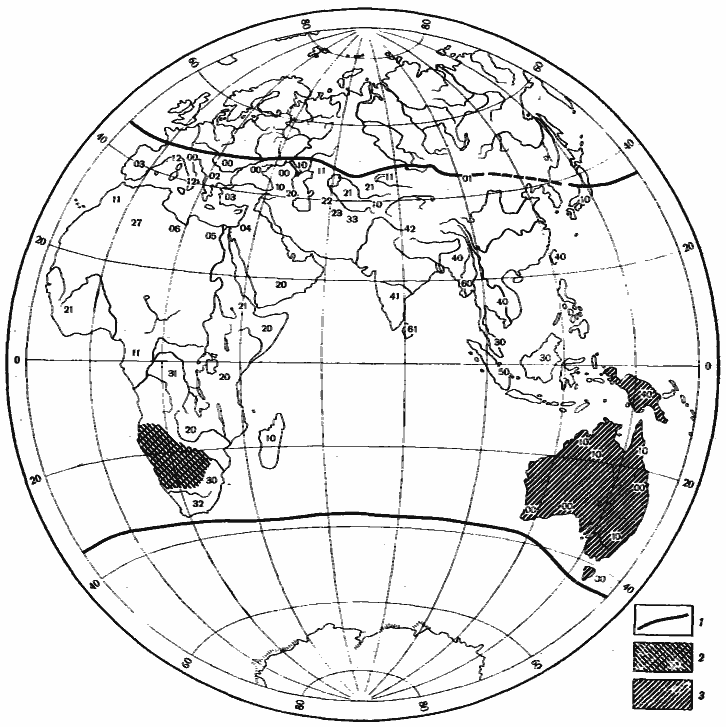
Рис. 12. Некоторые закономерности распространения Camponotus в Восточном полушарии
1 — границы распространения подрода Tenaemyrmex; индекс в пределах ареала подрода: первая цифра — число видов группы C. compressus—sylvaticus (в том числе C. xerxes и C. turcestanus), вторая — группы C. irritans—thraso (в том числе C. turcestanicus и C. fedtschenkoi); 2 — распространение подрода Myrmopsamma; 3 — распространение подрода Myrmophyma (по данным [Emery, 1920] с дополнениями)
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
обязательное существование только сообществами (семьями, общинами).
Довольно примитивная группа муравьев по своему строению и поведению. Рабочие мономорфны, самки по размерам близки к рабочим.
Небольшое наиболее архаичное подсемейство муравьев. Распространено по всему земному шару, преимущественно в тропиках. Насчитывает более 230 видов.
Всего 138 видов.
Включает устаревший: Monacis
организм, обитающий в древесном ярусе растительности.
Одно из крупных подсемейств, насчитывает около 1,5 тыс. видов, относящихся к 44 родам. Широко распространены на всем земном шаре, довольно обычны в тропиках и занимают доминирующее положение в умеренных широтах.
Всего 100 видов.
Всего 965 видов.
Всего 58 видов.
Всего 172 вида.
Включает сокращенный Novomessor.
Всего 65 видов.
Всего 62 вида.
Всего 11 видов.
Всего 13 видов.
Всего насчитывает 85 видов.
Всего 72 вида.
Myrmicinae — самое большое подсемейство муравьев, включающее более 2 тыс. видов, относящихся более чем к 150 родам.
Всего 68 видов
Всего 427 видов.
«медовые муравьи»
Всего 22 вида.
Всего 60 видов.
организм, обитающий в лесной подстилке.
организм, обитающий среди растительных или иных органических остатков, на поверхности почвы.
Всего 14 видов.
организм, обитающий на поверхности и в толще пород суши.
организм, питающийся семенами растений.
(лат. mandibule — челюсть, от mando — жую, грызу), первая пара челюстей у ракообразных, многоножек и насекомых. Синоним: жвала.
организм, питающийся исключительно другими животными
Всего 106 видов.
Всего 71 вид.
(зоонекрофаг), организм, питающийся мертвыми животными, гетеротроф
Всего 415 видов
Всего насчитывает 549 видов.
Всего 106 видов.
необратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.
Всего 24 вида.
Всего 4 вида.
Всего 6 видов.
специальное приспособление для переноски сухой почвы, распространенн у пустынных муравьев, образован длинными отстоящими волосками (аммохетами), часто имеющими форму латинской буквы J, расположен на нижней поверхности головы.
часть груди муравья, образоваванная слиянием заднегруди и первого сегмента брюшка.
у муравьев имеется три основные касты: самцы, самки и рабочие (бесплодные модифицированные самки).
рабочие с сильно раздутым брюшком; это каста, так называемых, "медовых бочек".
Всего 23 вида.
Всего 30 видов.
Всего 60 видов.
ксеробионт
Старое название: Proformica deserta
эволюционно сложившаяся, пространственно ограниченная, длительно самоподдерживающаяся, однородная экологическая система, в которой функционально взаимосвязаны живые организмы и окружающая их абиотическая среда.
 Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant
Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant Tetramorium caespitum (Linnaeus, 1758) — дерновый муравей, pavement ant
Tetramorium caespitum (Linnaeus, 1758) — дерновый муравей, pavement antСтарое название: T. indocile
животные, не имеющие позвоночника: простейшие, губки, кишечнополостные, черви, моллюски, членистоногие, иглокожие и др.; примерно 3/4 видов всех животных.
 Lasius alienus (Foerster, 1850) — бледноногий муравей, бледный садовый муравей, полевой муравей, cornfield ant
Lasius alienus (Foerster, 1850) — бледноногий муравей, бледный садовый муравей, полевой муравей, cornfield ant Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунок
Cataglyphis aenescens (Nylander, 1849) — степной бегунок, черный бегунокгигрофил, галобионт
Старое название: Cataglyphis flavigastra
Всего 19 видов.
Старое название: Leptothorax serviculus
Всего 48 видов.
организм, способный существовать в разнообразных условиях внешней среды
участок, используемый муравьями для добычи пищи.
 Formica cunicularia Latreille, 1798 — прыткий степной муравей
Formica cunicularia Latreille, 1798 — прыткий степной муравей Formica rufibarbis Fabricius, 1793 — краснощекий муравей, краснощекий муравей-минер
Formica rufibarbis Fabricius, 1793 — краснощекий муравей, краснощекий муравей-минерСтарое название: Anoplolepis longipes
Всего 17 видов.
итал. obligato, "обязательный", необходимый, существенный






