






Большие семьи могут выделять много тепла, влаги и углекислоты. Их проблема состоит в том, чтобы рассеивать CO2 и сохранять тепло и влагу соответственно своим потребностям: в прохладном климате надо сохранять тепло и избавляться от влаги, а в сухом — наоборот. Углекислота очень легко диффундирует сквозь почву и, несомненно, именно так удаляется из гнездовых холмиков, хотя из гнезд таких чувствительных к CO2 насекомых, как медоносные пчелы, этот газ приходится активно удалять путем проветривания. Тепло обычно сохраняется благодаря теплоизоляции, т. е. использованию пространств, в которых застаивается воздух, плохо проводящий тепло. Влага, если ее испарение не происходит достаточно быстро, удаляется при проветривании вместе с потоками воздуха. Тем не менее точно отрегулировать эти три переменные — задача весьма непростая.
Сохранение метаболического тепла в гнездах некоторых термитов хорошо установлено; например, у Nasutitermes exitiosus в обитаемых холмиках температура почти на 10° выше, чем в «мертвых» гнездах, а в камерах для расплода — всегда выше, чем поблизости в почве или воздухе (Holdaway, Gay, 1948); в Австралии температура в гнездах Coptotermes, расположенных в центральной части древесных стволов, колеблется от 33 до 38°C, что на 13—20° выше, чем в таких же деревьях, где нет термитов. В огромных и сложных термитниках Macrotermes к метаболическому теплу самих термитов добавляется тепло, выделяемое грибными садами. Сохранение этого тепла очевидно, и именно оно — причина того, что температура в участках гнезда, где много термитов, на 5° выше; температура в обитаемых холмиках равна в среднем 30°C с амплитудой колебаний в 4°. Далеко не все выделяемое тепло сохраняется, и возможно, что каналы и трубы, по которым циркулирует воздух, на самом деле охлаждают гнездо, а не собирают тепло. Воздух поднимается от грибных садов в верхушку холмика или в трубы и возвращается назад в полость или погреб. Это движение воздуха может быть конвекционным, но оно может также быть обусловлено ветром над трубами, высасывающим воздух наружу, или ветром на уровне почвы, задувающим его внутрь. Weir (1973) установил, что термиты Macrotermes устраивают отверстие на уровне земли; когда он закрывал это отверстие, стенки гнезда становились очень влажными и оно разваливалось. Weir высказал предположение, что сухие послеполуденные ветры охлаждают и осушают гнездо. Кристаллы солей, обнаруженные в погребах, могли выпасть из почвенных растворов после испарения влаги. Вентиляционный поток воздуха почти наверное способствует удалению углекислоты, хотя значительная ее часть может диффундировать через земляную корочку (рис:51).
Термиты приспосабливают структуру своих гнезд к данному местообитанию. В сухой местности им приходится добавлять к метаболической воде еще воду, добываемую из глубины почвы. Cubitermes exiguus выстилает свое гнездо большим количеством экскрементов, а некоторые виды Macrotermes в сухих местах устраивают гнезда на большей глубине. Macrotermes natalensis строит холмики в областях с прохладными зимами, например на плато в южной части Африки. Nasutitermes exitiosus в засушливых областях Австралии сохраняет воду так эффективно, что относительная влажность в его камерах редко бывает ниже 92%; поэтому полагают, что метаболической воды ему достаточно, так как наружная оболочка его холмиков толщиной 1—4 см состоит из глины, сцементированной экскрементами и, возможно, слюной (Lee, Wood, 1971, p. 63). В южных районах этот вид строит холмики на южных склонах, и обитатели гнезда в прохладную погоду собираются в его внутренней области, но зимней спячки в строгом смысле у них нет.
Лесные муравьи группы ![]() Formica rufa смогли прижиться в лесах умеренного пояса в значительной мере благодаря своим холмикам, построенным из растительных остатков. Некоторые муравьи, например F. exsectoides в северо-восточной части Северной Америки (Janzen, 1969, р. 152), уничтожают траву, кустарники и проростки деревьев на площади в несколько метров вокруг гнезда. F. ulkei вблизи Чикаго (41° с.ш.) в лесу из дуба и гикори в большинстве случаев начинает строить свои холмики на краю леса, и в дальнейшем они растут асимметрично — более широкая сторона обращена к свету. Их расположение определяется главным образом тем, откуда падают солнечные лучи, а не направлением магнитного поля Земли. Холмики непроницаемы для воды благодаря кровле толщиной 5 см из растительных остатков и земли; поэтому внутри гнезда круглый год бывает суше, чем в окружающей почве, и каждый холмик явно поглощает и накапливает тепло, так что на глубине 5 см на солнечном склоне температура в них достигает 47°C (Scherba, 1958, 1959, 1962).
Formica rufa смогли прижиться в лесах умеренного пояса в значительной мере благодаря своим холмикам, построенным из растительных остатков. Некоторые муравьи, например F. exsectoides в северо-восточной части Северной Америки (Janzen, 1969, р. 152), уничтожают траву, кустарники и проростки деревьев на площади в несколько метров вокруг гнезда. F. ulkei вблизи Чикаго (41° с.ш.) в лесу из дуба и гикори в большинстве случаев начинает строить свои холмики на краю леса, и в дальнейшем они растут асимметрично — более широкая сторона обращена к свету. Их расположение определяется главным образом тем, откуда падают солнечные лучи, а не направлением магнитного поля Земли. Холмики непроницаемы для воды благодаря кровле толщиной 5 см из растительных остатков и земли; поэтому внутри гнезда круглый год бывает суше, чем в окружающей почве, и каждый холмик явно поглощает и накапливает тепло, так что на глубине 5 см на солнечном склоне температура в них достигает 47°C (Scherba, 1958, 1959, 1962).
Холмики европейского вида Formica polyctena несколько крупнее и более рыхлые, а в их симметричном куполе, по-видимому, меньше минеральных веществ. Эти муравьи фуражируют на поверхности земли как в тени, так и на солнце, а также в кронах деревьев, но строят гнезда на краю леса, и если холмик оказывается затененным, то переходят на новое место. Изучая поведение фуражиров ![]() F. rufa у северной границы их ареала на Британских островах, Skinner (1980а) установил, что они избегают дождя и темноты, но в остальном интенсивность их фуражирования зависит от температуры.
F. rufa у северной границы их ареала на Британских островах, Skinner (1980а) установил, что они избегают дождя и темноты, но в остальном интенсивность их фуражирования зависит от температуры.
Изучая рассеивание тепла в двулетних гнездах Formica polyctena, Брандт (Brandt, 1980) установил, что в этих гнездах благодаря их структуре ежедневная волна тепла распространяется быстрее, чем в беспорядочных кучах материала, из которых они построены, или даже в кучах обычной почвы из тех же мест. В гнезде тепловая волна полностью затухает на глубине 20—30 см, а в почве или в куче гнездового материала — на глубине 10 см. Благодаря этому гнездо очень хорошо прогревается, сохраняет максимум тепла на ночь и в нем создаются прекрасные условия для созревания расплода. Брандт даже высказывает мнение, что гнездовой холмик по своим термическим характеристикам эквивалентен камню, и отмечает, что муравьи, в сущности, выигрывают, если кусты частично заслоняют гнездо от солнца, — это предотвращает чрезмерное нагревание поверхности (скажем, до 50° C). В гнездах больших размеров метаболическое тепло и тепло, выделяемое при аэробном разложении гнездового материала, может играть более важную роль. Coenen-Stass et al. (1980) нашли, что хотя все обитатели муравейника создают больше тепла, чем равное по массе количество гнездового материала, однако ввиду того, что этого материала гораздо больше, его теплопродукция тоже больше; 82 кг гнездового материала давали в 7 с лишним раз больше тепла, чем 1 кг муравьев. В целом гнездо давало 16,21 Вт, а его население — 2,62 Вт, что в сумме составляет около 19 Вт (рис:52).
Остается рассмотреть роль, которую играет во всем этом поведение рабочих; они выбирают места, обогреваемые солнцем, обрабатывают материал, идущий на построение гнезда, открывают и закрывают входные отверстия, выходят на солнце и снова прячутся. Но можно ли считать, что муравьи собирают для приготовления компоста наилучший материал? Действительно ли хвоинки, почечные чешуйки и веточки наиболее пригодны для этого или просто их легче всего собирать? Лесные муравьи строят холмики, которые поглощают и сохраняют тепло солнечных лучей и, медленно разлагаясь, дают довольно значительное количество «фонового» тепла. Кроме того, муравьи сохраняют собственное метаболическое тепло, однако все еще остается нерешенным вопрос: увеличивают ли они свою теплопродукцию при действительном или ожидаемом понижении температуры в гнезде?
Относительное постоянство гнезд у ос и пчел связано с тем, что их микроклимат в большей мере зависит от выбора места и от способности этих насекомых корректировать отклонения от оптимума своим поведением. Например, если у гнезда нет оболочки, то осы или пчелы прикрывают его от солнца или дождя своими телами; если же нужно создать сквозняк или распылить воду, чтобы она испарялась, то они машут крыльями.
Гнезда Polistinae, подвешенные на ножке, часто бывают расположены под каким-нибудь укрытием, но если гнездо ничем не защищено, то осы сидят на нем, «свесив хвосты», так что дождевая вода стекает по их крыльям и брюшку. Оболочки, если они есть, защищают расплод от прямых солнечных лучей, а регуляция температуры, влажности и концентрации углекислоты в гнезде достигается вибрацией крыльев, хотя в результате мышечных сокращений температура тела осы может при этом повыситься на 3° (Evans, West-Eberhard, 1973). В сухом климате осы Polistes собирают воду, чтобы охлаждать свои гнезда.
В вырытых в земле норках пчел Halictidae надлежащий микроклимат обеспечивается, по-видимому, выбором места для норки и ее глубиной, как и у одиночных видов. Иногда, однако, эти пчелы производят вентиляцию, располагаясь у входа в норку (Sakagami, Michener; 1962). Пчелы Meliponinae либо избегают неподходящих мест, либо регулируют микроклимат в гнезде. К первому типу относится Leurotrigona miilleri, совершенно неспособная регулировать среду в гнезде, но обитающая обычно в гнездах термитов; Trigona (Apotrigona) nebulata аналогичным образом устраивается в расположенных на деревьях шарообразных гнездах одного из видов Nasutitermes диаметром 30—50 см. Пчелы, живущие в норах, которые они выстилают церуменом, если их поселить в искусственные ульи, часто оказываются неспособными регулировать температуру (Darchen, 1973). Meliponinae в саванне нередко строят гнезда в заброшенных холмиках Macrotermes или Trinervitermes. Старые холмики сначала разрушаются позвоночными и вновь заселяются другими термитами (например, Amitermes, Ancistrotermes) или муравьями (представителями Camponotus, Crematogaster или других родов); позднее в центре этого комплекса поселяются пчелы, устраивающие длинную извилистую трубку, ведущую от их гнезда наружу.
В отличие от этих форм другие Meliponini способны очень эффективно регулировать температуру. В Бразилии Trigona spinifer поддерживает в своих гнездах температуру 33—36°C при температурах окружающей среды от 8 до 30°C (Zucchi, Sacagami, 1972). Так же обстоит дело у Dactylurina, которая поселяется в густых зарослях растений, где она строит шарообразные гнезда с толстой (9 см) пенистой обшивкой из галерей и тонкой смолистой оболочкой толщиной всего 0,5 см. Пчелы вентилируют гнездо, махая крыльями; для более эффективной вентиляции они проделывают отверстие на стороне, расположенной против входной трубки, или же делают несколько мелких отверстий в оболочке. Трудно поверить, каким громким может быть шум их вибрирующих крыльев в жаркий полдень. Маленькие пчелы Hypotrigona, живущие в полостях веток, вентилируют гнездо, выстроившись в ряды головами к центру гнезда (Darchen, 1973). Несколько видов безжалых пчел благодаря своей способности регулировать температуру смогли проникнуть в Южную Америку до областей с холодными зимами; здесь они зимой не размножаются, но и не делают никаких специальных приготовлений к зимовке.
Открытое гнездо обитающей в тропиках Азии пчелы Apis dorsata бывает покрыто тремя или более слоями пчел, которые висят на нем брюшком вниз, зацепившись коготками. По этой «завесе» очень хорошо стекает дождевая вода, а кроме того, она защищает расплод от солнца и способствует сохранению тепла. Если температура падает ниже 30°C, пчелы сбиваются в кучу, чтобы уменьшить потерю тепла; если же становится слишком жарко, они начинают вибрировать крыльями или даже собирают воду, которая затем испаряется. Все это позволяет им поддерживать в соте температуру 30—31°C при внешних температурах от 20 до 34°C (Morse, Laigo, 1969; цит. по Michener, 1974). То же самое проделывает A. florea в восточной Африке (20° с.ш.), удерживая температуру в гнезде на уровне 33—37°C при температурах среды от 18 до 33°C (Free, Williams, 1979). Ни тому, ни другому виду Apis, по-видимому, нет нужды генерировать тепло днем; для поддержания нужной температуры им достаточно теплого климата, нормального метаболизма и теплоизоляции, создаваемой слоем пчел. В равной мере это относится и к муравьям-кочевникам Eciton hamatum (рис:53).
Такая реакция, направленная на обогревание гнезда, могла возникнуть из инкубационного поведения, когда взрослые особи путем тесного контакта своих тел с расплодом создают для него подходящий микроклимат. На это указывают данные о тех галиктах, которые строят грозди ячеек под землей. Шершни и осы прижимаются к расплоду и поочередно втягивают и растягивают брюшко, распространяя тепло, вырабатываемое в груди, и одновременно усиливая газообмен. Они не инкубируют пустые коконы, но будут инкубировать кусочки фильтровальной бумаги, смоченные экстрактом из куколок, — несомненный признак химического опознавания (Ishay, 1972).
Молодая самка осы Vespula плотно прижимается к своим первым ячейкам с расплодом, что вполне может повысить их температуру, а поскольку ячейки покрыты оболочкой, то и температуру воздушного пространства. Однако наивысшего искусства в инкубировании расплода достигает Bombus. Самка, начинающая строить гнездо, делает в первом личиночнике неглубокую канавку, в которую плотно входит ее тело (Alford, 1975). Запас меда в медовой чаше, расположенной в пределах досягаемости, служит «топливом» для процесса дыхания, и температура расплода становится значительно выше окружающей температуры (Michener, 1974). Heinrich (1974) сумел показать, что самка в состоянии поддерживать температуру груди на уровне 35—38°C, а температуру брюшка на уровне 31—36°C. Это дает ей возможность согревать расплод до 25°C при температуре среды 5°C и до 33°C при температуре среды 30°C, т. е. она явно способна к терморегуляции. С понижением температуры среды потребление самкой кислорода повышается; очевидно, она оценивает степень отклонения от оптимума и соответственно повышает свой метаболизм за счет активности летательной мускулатуры, т. е. действует подобно простому гомеостату. Успех этой реакции зависит от ее быстроты и от термоизоляции гнезда; в более благоприятном положении окажутся, конечно, те самки, которые выбрали (или которым посчастливилось найти) подходящее место для гнезда. Должно ли оно находиться на солнце или в тени? В одном случае экономится топливо, в другом обеспечивается более постоянный режим. По мере увеличения числа рабочих особей терморегуляция улучшается, особенно при постройке навеса. При этом температура в гнезде поддерживается на уровне 30°C с колебаниями в диапазоне менее чем 3°, и остается лишь предотвращать перегрев, вдувая в гнездо прохладный воздух; собирать воду при этом не приходится.
Vespinae, обитающие в умеренном климате в подземных гнездах, выстланных бумагой, тоже превосходно регулируют температуру гнезда, особенно летом, в период наибольшей численности населения; зимой они живут в одиночку, а их самки диапаузируют, подобно большинству насекомых. Оптимальная температура для их расплода около 30—33°C, и они могут поддерживать ее даже при температуре среды 10°C; для охлаждения гнезда применяется вентиляция, а в случае необходимости — сбор воды и ее испарение (Edwards, 1980).
У Apis mellifera и близкого к ней вида A. cerana кондиционирование среды в гнезде приближается к совершенству. Эти виды распространены во всех областях с умеренным климатом, где они могут жить без помощи человека в запечатанных полостях с термоизоляцией, так как запасают очень много меда и медленно используют его на протяжении всей зимы. Они не должны допускать слишком сильного снижения температуры в гнезде: при 8°C рабочие особи уже гибнут. Подготовка к зиме начинается с укорочением дня, задолго до наступления зимних холодов; матка прекращает откладку яиц, а молодые личинки уничтожаются, так что зимуют только взрослые особи.
Когда температура падает ниже 18°C, пчелы сбиваются в клуб, наружный слой которого образуют плотно прижавшиеся друг к другу неподвижные пчелы, создающие термоизоляцию, тогда как особи, находящиеся внутри клуба, активно двигаются и «дрожат», чтобы продуцировать тепло (летательные мышцы могут вибрировать, не приводя в движение крылья). Эти две группы пчел редко меняются местами. Внутри клуба может поддерживаться температура выше 20°C при температуре среды, близкой к нулю. Любопытно, что соты почти не мешают формированию клуба и движению в нем пчел; только пчелам Apia cerana приходится проделывать в сотах отверстия, чтобы стали возможными поперечные передвижения. Поглощаемый в этот период сироп очень густой, но пчелы поддерживают нужную влажность за счет метаболической воды. Они удерживают в организме свои (жидкие) экскременты и вылетают наружу лишь весной, с возвращением яркого солнца и тепла.
Весной пчелы доводят температуру в небольшой части гнезда с расплодом до 35°C. Возможно, они больше реагируют на длину дня, чем на внешнюю температуру, которая сильно варьирует. Летом, когда численность населения в улье велика, температура расплода может регулироваться с точностью почти до 0,5° (Biidel, 1968). Такой стабильности способствует большая масса расплода и меда, которые весьма эффективно сохраняют тепло, хотя слабо продуцируют его (в этом отношении даже вклад расплода пренебрежимо мал). Влажность обычно бывает достаточной; пчелы вентилируют гнездо не только для того, чтобы удалить из него нагретый воздух, но и для того, чтобы впустить сухой воздух, необходимый для испарения воды из нектара во время созревания меда, что тоже способствует охлаждению улья. Все это приводит к тому, что пчелы лучше чувствуют себя в сухой атмосфере, чем во влажной (Michener, 1974). При исключительно жаркой и сухой погоде пчелы собирают воду и распыляют ее по сотам.
Lensky (1963, цит. по Darchen, 1973) показал, что Apis mellifera ligustica (итальянская раса) в Израиле может поддерживать в гнезде с расплодом температуру 34—38°C при наружных температурах от 22 до 50°C. Такая терморегуляция, по-видимому, недоступна одному из африканских подвидов — A. mellifera adansonii, который старается избежать перегрева, поселяясь в разного рода пустотах между корнями деревьев, ограничивая численность семей путем частого роения и мигрируя в новые места (Darchen, 1973). Пчелы реагируют таким образом не только на перегрев во время сухого сезона, но и на недостаток пищи в период дождей (как, например, A. cerana indica) (Woyke, 1976) или на нападение млекопитающих, муравьев или других членистоногих, например пчелиной огневки (Fletcher, 1975). Готовясь к переселению, пчелы выбрасывают личинок из запечатанных ячеек, прекращают откладку яиц и поедают запасы меда. Вероятность переселения выше для семей, которые недавно роились, и некоторые виды склонны к этому больше, чем другие (Winston et al., 1979). Переселение явно представляет собой необычную форму миграции, характерной для A. florea и A. dorsata.
Во время вентиляции из гнезда, конечно, удаляется углекислота, хотя пчелы способны выдержать довольно высокое ее содержание в воздухе, вплоть до 3%. При повышении концентрации CO2 Apis mellifera начинает вентилировать гнездо, даже если температура ниже оптимальной; в очень больших семьях содержание CO2 регулируется более эффективно, чем в маленьких (Seeley, 1974).
Подведем итоги. Муравьи выбирают подходящее место и строят многокамерное гнездо, где представлено широкое разнообразие микроклиматов, по которым можно распределить расплод. В прохладном климате они используют для накопления тепла такие природные объекты, как плоские камни, и строят на освещаемых солнцем местах холмики, поглощающие и сохраняющие тепло; есть виды, способные даже использовать тепло, выделяющееся при аэробном разложении растительных материалов, которые они собирают в кучу, а другие виды, может быть, способны генерировать метаболическое тепло и в ответ на понижение температуры среды. В жарком сухом климате муравьи избегают солнца, добывают воду из глубоких слоев почвы и из растений или собирают росу. Термиты менее подвижны, чем муравьи, так как у них репродуктивные камеры гнезда часто бывают жестко фиксированными, но они тщательно выбирают места для постройки гнезда, покрывают его оболочкой, по которой скатывается вода, и регулируют потоки воздуха, изменяя величину отверстий, ведущих в ходы и полости.
Холмики имеют теплоизоляцию, что позволяет термитам сохранять или рассеивать метаболическое тепло в зависимости от обстоятельств. Осы и пчелы могут инкубировать свой расплод и покрывать гнезда теплоизолирующими материалами; они могут также при колебаниях температуры покрывать гнездо плотной живой оболочкой из собственных тел, продуцировать тепло, повышая интенсивность своего метаболизма при нехватке тепла, или же рассеивать тепло (а также газообразные отходы и водяные пары) путем вентиляции и испарения воды при угрозе перегрева.
Медоносные пчелы способны очень тонко регулировать температуру летом и не допускать охлаждения гнезда зимой. Эта активная регуляция микроклимата требует расхода энергии, но необходима из-за неподвижности расплода. После специальной подготовки, проводимой летом, целые семьи могут переходить на новое место, захватив с собой большую часть запасов. Пчелы и осы, а в меньшей степени и муравьи переселяются, вероятно, в ответ на серьезную угрозу климатического или биотического характера, и если это случается регулярно, то переселения могут перейти в миграции. Дальнейшие подробности см. Seeley, Heinrich (1981).
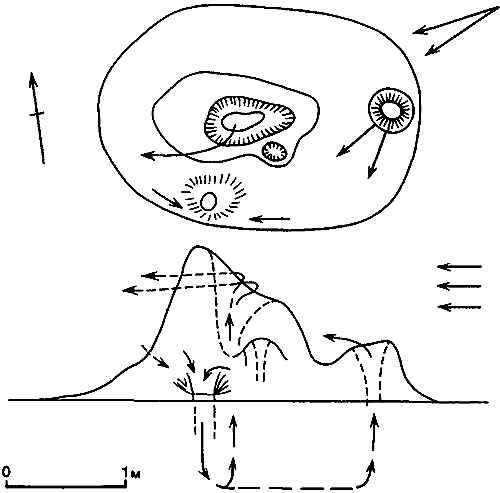
Рис. 51. Направление ветра и циркуляция воздуха в гнездовом холмике Macrotermes subhyalinus. (Weir, 1973.)
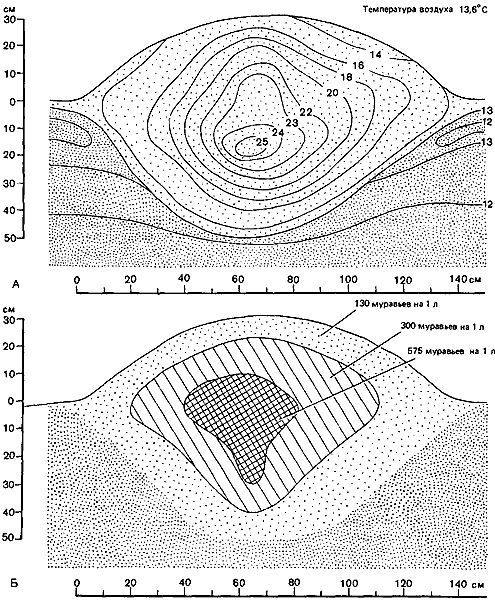
Рис. 52. Распределение температуры (°C) в гнездовом холмике Formica polyctena (А) и плотность муравьев в это же самое время (Б) (Coenen-Stass et al., 1980.)
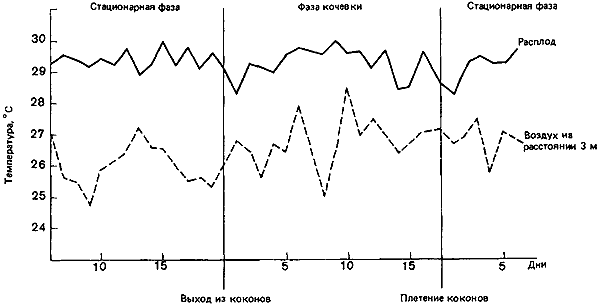
Рис. 53. Температура внутри и вокруг клуба Eciton hamatum, измерявшаяся ежедневно на протяжении полного цикла их активности. (Schneirla, 1971.)
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
надсемейство летающих насекомых отряда перепончатокрылых, родственное осам и муравьям. Существует около 20 тысяч видов пчёл. Распространены на всех континентах, кроме Антарктиды. Пчёлы питаются нектаром и пыльцой, используя нектар главным образом в качестве источника энергии, а пыльцу для получения белков и других питательных веществ.
отряд общественных насекомых. Общины, разделенные на касты, состоят из крылатых и бескрылых особей. Строят подземные и наземные (до 15 м высотой) гнезда (термитники). Ок. 2600 видов, главным образом в тропиках; в России 2 вида: один в районе Сочи, второй, вероятно, завезенный из Японии или Китая, во Владивостоке. Многоядны. Разрушают древесину, портят бумагу, кожу, сельскохозяйственные продукты и др. Термиты, обитающие в почве, полезны как почвообразователи.
 Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравеймуравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
сборное название насекомых подотряда жалящих перепончатокрылых (кроме пчел и муравьев). Длина до 40 мм (шершни). Ок. 23 тыс. видов. Распространены широко. Опылители растений. Личинки некоторых ос паразиты других насекомых.
Всего 965 видов.
Всего 427 видов.
(лат. Vespa) род крупных общественных ос, относящихся к семейству Vespidae.
земляные пчёлы (лат. Bombus) — род перепончатокрылых насекомых, во многих отношениях близкий медоносным пчёлам.






