






У общественных пчел Halictidae и Bombini (Apidae) сбор пищи производится отдельными особями, которые не кооперируются друг с другом для выполнения этой задачи (Michener, 1974). Это не означает, что рабочие особи, достигшие достаточно зрелого возраста и достаточно крупные, не могут получить полезную информацию от других особей, прилетающих обратно в гнездо: особый звук, издаваемый при полете возвращающимися шмелями-фуражирами, отличает даже человек (Schneider, 1972, цит. по Michener, 1974). Вряд ли можно также сомневаться в способности пчел распознавать цветки, послужившие источником корма, по пыльце, приставшей к фуражирам, или же по вкусу отрыгиваемой ими жидкости. В результате насекомые могут сосредоточить свои усилия на нескольких из большого числа возможных кормовых растений, растущих но соседству, как сообщает Free (1970) относительно Bombus lucorum.
Осы тоже способны информировать членов своей семьи о наличии корма; возможно, в этом участвует обоняние, так как известна необычайная чувствительность ос к химическим веществам (Holldobler, 1977). Вылеты и возвращения в гнездо как шмелей, так и ос распределены во времени не случайным образом, однако анализ данных о B. affinis, проведенный Плоурайтом (Plowright, 1979), позволяет предполагать, что причина этого — внешние изменения, связанные с погодой, а не поведение других особей.
В природе виды Bombus используют цветки, глубина венчика которых соответствует длине их хоботка, хотя эта корреляция никогда не бывает очень тесной, так как и насекомым, и цветкам свойственна большая внутривидовая изменчивость по величине, а нектар удается иногда достать даже при длинной трубочке венчика. Например, синяк обыкновенный (Echium vulgare) вырабатывает нектар со скоростью, зависящей от температуры воздуха, и пчела, у которой хоботок относительно короткий, может добраться до нектара только в сырую погоду, когда он более обилен, хотя и менее концентрирован.
Наблюдалось, что медоносные пчелы собирали нектар даже в сухую погоду на одном месте, где у растений были необычно короткие венчики (Corbet et al., 1979), а к клеверу луговому (Trifolium pratense) они подлетают только тогда, когда нектар поднимается в трубке венчика достаточно высоко, чтобы они могли до него дотянуться (Free, 1970). У многих видов Bombus (например, у B. hortorum) хоботки очень длинные, что позволяет им быть единственными потребителями нектара растений с глубоко расположенными нектарниками.
Вероятно, пчел разных видов привлекают цветки различного типа, соответствующие длине их хоботков, однако мало что известно о том, какими указаниями они при этом пользуются. До того как пчела вылетит из гнезда, она может располагать сведениями лишь о вкусе и запахе, но не об окраске и форме цветков; поэтому возможно, что в самом начале ее деятельности как сборщицы главную роль играют вкусовые и обонятельные реакции. Между тем окраска тоже имеет значение при выборе цветка, так как цветки с короткими венчиками чаще бывают белыми или желтыми, а цветки с длинными венчиками — синими или пурпурными (Knuth, 1906—1909, цит. по Proctor, Yeo, 1973). Три вида Bombus чаще выбирали желтые или оранжевые искусственные цветы, чем синие или пурпурные, а реже всего — красные или белые (Brian A. D., 1967). Кроме того, шмелей легче приучить к синим, чем к белым искусственным цветам; в опытах с белыми цветами вознаграждение должно быть больше, а приобретенный навык забывается быстрее (Heinrich et al., 1977).

Рис. 16. Число шмелей, собирающих нектар из цветков липы (Tilia) в зависимости от концентрации сахара в нектаре. (Corbet et al., 1979.)
Возможно, однако, что имеет значение и форма, так как у цветков с длинными венчиками она чаще бывает неправильной или двусторонне-симметричной; это обычно одиночные цветки, свисающие вниз, так что пчелам приходится подлетать к ним снизу. В отличие от этого цветки с короткими венчиками часто образуют соцветие, например головку, как у Compositae; такие цветки привлекают к себе Bombus lapidarius (Prys-Jones, 1982). Некоторые шмели стараются сесть на цветок, на котором уже сидят другие пчелы (Brian A. D., 1957); конечно, такое поведение, хотя оно, возможно, в основном носит агрессивный характер, могло бы сильно сокращать поиски, приводя фуражиров к подходящим кормовым растениям.
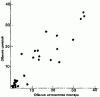
Шмели научаются узнавать цветки, которые им подходят, и приобретают опыт, как лучше всего подлетать к ним и проникать в них. Heinrich (1979) выпускал молодых насекомых на природный луг, защищенный сетками от других диких пчел; шмели перепробовали пять видов растений, прежде чем остановились на бальзамине (Impatiens) — растении с двусторонне-симметричными цветками и глубоко расположенными нектарниками. Но первые посещения даже этого цветка были неудачными: шмели садились на верхнюю часть венчика, а не на губу, и только после 60 попыток они приобрели достаточный навык и могли наполнить свой зоб, посетив 20 цветков за 6,2 мин. В отличие от этого для посещения радиально-симметричных головчатых соцветий астры (Aster) и золотой розги (Solidago) они не нуждались в опыте, но и пользу извлекали при этом минимальную.
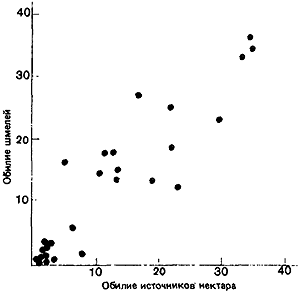
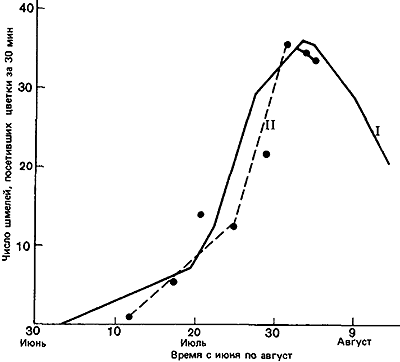
Важное значение запасов нектара в цветке выявилось, когда всех пчел выпустили сразу: запасы нектара в цветках бальзамина уменьшились до одной четверти, в клевере луговом и астрах — наполовину, а в золотой розге вовсе не уменьшились. Таким образом, конкуренция заставила некоторых насекомых отдавать предпочтение клеверу по сравнению с бальзамином. Далее, если клевер ползучий (Trifolium repens) и горошек мышиный (Vicia cracca) на определенном участке были защищены от пчел сеткой и накопили запасы нектара, то Bombus vagans будет позднее оказывать им больше внимания, чем цветкам тех же видов в других местах (Morse, 1980). Шмели обычно предпочитают более концентрированный нектар (рис:16), но вязкость может затруднить шмелям с их длинным хоботком высасывание слишком густого нектара, так что имеется некий оптимум концентрации, при котором «чистая прибыль» в отношении энергии будет для насекомого максимальной (Prys-Jones, 1982). Pleasants (1981) показал, как изменяется численность шмелей на протяжении сезона в соответствии с обилием цветов (рис:17 и рис:18; данные для одного района Скалистых гор в шт. Колорадо).
Рис. 18. Сезонные изменения количества нектара (I) и численности шмелей Bombus bifarus (II), которые собирали нектар во время 30-минутных наблюдений. (Pleasants, 1981.)
До сих пор мы рассматривали сбор нектара. Сомнительно, чтобы шмели с их длинными хоботками когда-либо собирали пыльцу без нектара; они принадлежат к очень специализированной ассоциации цветок—насекомое. Другие пчелы не таковы — они посещают и цветки, дающие только пыльцу, например мак (Papaver). А на таких растениях, как подсолнечник (Helianthus annuus), дающий и нектар, и пыльцу, насекомые, если они собирают пыльцу, посещают лишь те цветки соцветия, у которых только что лопнули пыльники. Пыльца высыпается, после того как насекомое потрясет пыльники подсолнечника, а у цветков других типов пчелы «выдаивают» или откусывают пыльцу, либо собирают, когда она уже высыпалась (Eickwort, Ginsberg, 1980).
Сборщицы нектара могут посетить те же места и оказаться усыпанными пыльцой, которую они перенесут на более старые цветки в том же соцветии подсолнечника, ползая по нему. Однако нередко они счищают пыльцу своими щеточками и выбрасывают ее; но не потому, что пыльца им не подходит, — просто в данный момент она их не интересует. Несмотря на такое поведение, они могут превосходить других насекомых как опылители (Free, 1970). У пчел с высокоразвитым общественным образом жизни сбор пыльцы и нектара нередко происходит во время разных вылетов при посещении разных цветков, а сборщицы пыльцы, готовясь к походу, пьют запасенный в улье мед. Такие виды могут также собирать воду для разбавления запасенного меда; количества главных пищевых продуктов (нектара, воды, пыльцы), которые нужно собрать, несомненно, регулируются путем обмена информацией внутри гнезда.
Опыление было бы более эффективным, если бы данное насекомое в течение некоторого времени посещало растения одного вида, вместо того чтобы беспорядочно перелетать от одного вида к другому. Удалось ли растениям принудить насекомых к такой тактике? В случае медоносной пчелы почти удалось, так как в ее сборах смешанная пыльца составляет всего 6%. С Bombus дело обстоит хуже: у B. lucorum такой пыльцы уже 34%, а у B. pascuorum — 63% (Free, 1970). Это может помочь перекрестному опылению травянистых растений и мелких кустарников, но не деревьев, так как насекомое скорее перейдет с одной ветки данного дерева на другую, чем на какое-то новое дерево. Чтобы это не приводило к самоопылению, у многих деревьев в процессе эволюции образовались отдельные мужские и женские цветки (Maynard Smith, 1978а, р. 136). Пыльца деревьев-опылителей, посаженных в плодовом саду, все же переносится на соседние деревья; предполагают, что этот перенос происходит в результате последовательных посещений шмелей, так как пыльца, приставшая к их волоскам, сохраняет жизнеспособность несколько часов (Free, Williams, 1972).
Содержание сахара в нектаре и количество пыльцы варьируют в течение дня и зависят от внутренних ритмов раскрывания цветков, освобождения пыльцы и чувствительности рыльца, а все эти факторы изменяются в зависимости от погодных условий. Например, у бобов (Vicia faba) цветок в первый день раскрывается для того, чтобы выбросить пыльцу, во второй день — чтобы воспринять ее (рыльце созрело), а на третий день он вянет и уже не привлекает шмелей. У этой группы растений соцветие обычно представляет собой побег, на котором вместо листьев растут цветки; у него есть верхушечная точка роста, производящая молодые бутоны, каждый из которых приобретает окраску, раскрывается, источает аромат, сбрасывает пыльцу, образует рыльце, готовое к ее приему, и наконец, увядает и становится незаметным.
Такая программа обеспечивает шмелям возможность сесть у основания соцветия и нанести свежую пыльцу на зрелые рыльца, а затем перейти или перелететь (в зависимости от расстояния между цветками) на самый верхний из раскрывшихся цветков; после этого шмель доставляет новую порцию свежей пыльцы самому старому, наиболее низко расположенному цветку на соседнем растении, и так далее (Free, 1970). Пайк (Руке, 1978, 1979) изучал, как Bombus appositus кормится на аконите (Aconitum columbianum), одновременно опыляя его, и установил, что этот шмель не поднимается систематически по возрастной спирали. Он, летая между цветками, чаще всего (в 90% случаев) переходит на ближайшего соседа сверху, а не на следующий более молодой цветок по спирали. Это сокращает летное время, хотя ведет к тому, что некоторые из цветков данной спирали будут пропущены; Пайк сформулировал следующее правило поведения шмелей: «лети на любой из самых ближайших цветков, расположенных выше, но не на тот, который посетил только что...».
[i107_tn.gif]
[i108_tn.gif]
Конечно, наблюдались и нарушения этого правила: один раз из пяти шмель перемещается вниз, а не вверх, а два раза из трех он покидает соцветие между основанием и верхушкой, т. е. посещает самый молодой цветок реже чем один раз из пяти. Тем не менее эта система работает, и в каждом отдельном вылете вторичные посещения данного цветка составляют лишь 1,2% общего числа посещений. Пайк указывает, что план, по которому следует переходить на ближайший цветок, а не на следующий цветок в спирали, хорош до тех пор, пока колоски (или растения) находятся не слишком далеко друг от друга. Если же расстояния между ними велики, то лучше сосредоточиться на одном соцветии и посетить все цветки без пропусков.
Другой случай, когда выгоден более поспешный сбор нектара, — это посещение цветков губовника линейного (Chilopsis liniaris) шмелями Bombus sonorus. В каждом цветке нектар, стекая по пяти желобкам, образует большую каплю. По утрам самки облетают цветки и высасывают эту каплю, получая 1,7 мкл нектара за 2 с; затем они возвращаются и опустошают желобки, получая за 5 с только 2,4 мкл нектара. Уитхэм (Whitham, 1977) вычислил, что при этом шмели получают на 25% больше нектара, чем, если бы они высасывали его сразу за одно посещение цветка; это выгодно и растению, так как число посещений цветка оказывается вдвое больше.
Более крупные шмели не только обладают большей силой, но и быстрее перемещаются от соцветия к соцветию, хотя и не от цветка к цветку; однако затраты энергии при полете возрастают пропорционально весу тела, так что крупный шмель расходует больше энергии, чем маленький. Оптимальные (наиболее экономичные) размеры для Bombus appositus, кормящегося на аконите, соответствуют весу 0,24 г, что близко к фактическому среднему весу, равному 0,28 г (Руке, 1978). Модель, которую построил Пайк, предсказывает также, что чем больше нектара в каждом цветке, тем крупнее должен быть посещающий его шмель; и в самом деле, B. appositus и B. kirbyellus кормятся на дельфиниуме и аконите, выделяющих в сутки по 2—4 мкл нектара, тогда как B. bifarius и B. sylvicola посещают соцветия сложноцветных, отдельные цветочки которых имеют венчики с короткой трубкой, причем одновременно раскрыты бывают лишь несколько цветочков (Proctor, Yeo, 1973).
В другой модели, описывающей сбор нектара шмелем, рассматриваются три состояния — шмель в гнезде, шмель в полете и шмель в цветке — и предполагается, что насекомое возвращается в гнездо после наполнения зоба (Oster, Wilson, 1978). Время, проведенное в цветке, зависит от строения цветка и количества содержащегося в нем нектара, а временной интервал между посещениями двух цветков — от их взаимного расположения (в частности, от расстояния между ними) и от скорости полета шмеля с учетом расхода энергии на взлет, посадку и на самый полет. При этом крупные шмели оказываются более эффективными при фуражировке в крупных цветках с большим количеством нектара, если только цветки расположены достаточно далеко друг от друга. Авторы предлагают формулу, выражающую прирост энергии на одного фуражира, который возрастает с увеличением плотности расположения цветков до известного предела, и предсказывающую оптимальные размеры фуражира при различной величине цветков. В общем более крупные шмели оказываются более эффективными фуражирами, но, конечно, для создания их самих требуются большие затраты.
Идея об оптимальной фуражировке стимулировала изучение факторов, влияющих на эксплуатацию данного «пятна» корма. Фуражир должен привести все «пятна», находящиеся на одинаковых расстояниях от его гнезда, к единому уровню в смысле поставки нектара; этот уровень соответствует ожидаемой скорости всасывания нектара, оцениваемой путем обобщения прошлого опыта (Charnov, цит. по Hassell, Southwood, 1978). Чем дальше от гнезда находится «пятно» корма, тем полнее оно должно быть использовано. Ходжес (Hodges, 1981), наблюдая за самками Bombus appositus, собирающими нектар на Delphinium nelsoni в Скалистых горах в шт. Колорадо, получил данные, согласующиеся с моделью Чарнова. Количество нектара в цветках одного соцветия (одного «пятна») было примерно одинаковым, так что шмелю достаточно было обследовать один цветок, чтобы решить, стоит ли продолжать сбор или лучше перейти на другое соцветие. Когда количество нектара было экспериментально увеличено, шмели стали дольше задерживаться на каждом соцветии и посещать все его цветки (их обычно около трех).
Несмотря на превосходную систему обмена информацией, существующую у медоносных пчел, иногда отдельные их особи трудятся поодиночке. Поведение таких пчел при сборе нектара изучали Butler et al. (1943), а при сборе пыльцы — Ribbands (1949). В обоих случаях отдельные пчелы были связаны с определенной группой цветков на небольшом участке, даже если данное растение занимало большую площадь. Пчелы посещали эти цветки до тех пор, пока запасы нектара или пыльцы оставались достаточными, но покидали их еще до того, как эти запасы иссякали полностью, хотя другие пчелиные все еще использовали их. Очевидно, данная пчела перепробовала другие растения и «... степень ее склонности к перемене определяется тем, что она помнит о прежних урожаях...» (Ribbands, 1949, р. 63). Ribbands подчеркивает, что пчелам свойственны «непрерывные попытки делать выбор...» Короче говоря, они непрерывно обследуют источники пищи и используют наилучший из тех, которые им доступны. Weaver (1957) предполагает, что они начинают исследовательскую деятельность после того, как насытятся на главном источнике нектара.
Существует еще одно своеобразное явление: некоторые шмели собирают нектар, прокусывая трубку венчика еще до того, как раскроется цветок, а насекомые, не способные к этому, используют дырки, проделанные другими видами (или даже птицами). В Европе таким разбойным нападениям подвергаются, по крайней мере, 300 видов растений (Brian A. D., 1954). По мнению некоторых исследователей (Heinrich, Raven, 1972), воровство нектара просто заставляет шмелей-опылителей с длинным хоботком посещать большее число цветков, тем самым способствуя аутбридингу. Такой «незаконный» сбор нектара означает, что цветок одного растения, например бобов, может обеспечить энергией опыление цветков другого, неродственного вида, например мака, который не вырабатывает нектара. Это действует эффективно и, возможно, обусловлено отчасти тем флористическим хаосом, который по воле человека сменил прежние стабильные или, во всяком случае, медленно изменяющиеся экосистемы. Старомодные шмели с длинными хоботками, получающие всю свою пищу от растений одного вида, как, например, Bombus hortorum — от окопника лекарственного (Symphytum officinale) (Prys-Jones, 1982), быть может, в очень близком будущем сохранятся только в заповедниках.
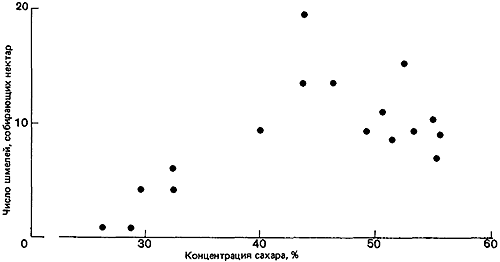
Рис. 16. Число шмелей, собирающих нектар из цветков липы (Tilia) в зависимости от концентрации сахара в нектаре. (Corbet et al., 1979.)
надсемейство летающих насекомых отряда перепончатокрылых, родственное осам и муравьям. Существует около 20 тысяч видов пчёл. Распространены на всех континентах, кроме Антарктиды. Пчёлы питаются нектаром и пыльцой, используя нектар главным образом в качестве источника энергии, а пыльцу для получения белков и других питательных веществ.
земляные пчёлы (лат. Bombus) — род перепончатокрылых насекомых, во многих отношениях близкий медоносным пчёлам.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
разноцветные
 (белые, желтые,оранжевые, коричневые) комочки неправильной формы, ~2-3мм диаметром, обножка это то что пчелы приносят на лапках. Слегка сладковатая на вкус с ярким цветочным привкусом. Пыльца — природный концентрат, содержащий белки, все известные витамины, ценные минеральные вещества, по общему аминокислотному составу обножка близка к другим богатым белком пищевым продуктам — мясу, молоку, яйцам.
(белые, желтые,оранжевые, коричневые) комочки неправильной формы, ~2-3мм диаметром, обножка это то что пчелы приносят на лапках. Слегка сладковатая на вкус с ярким цветочным привкусом. Пыльца — природный концентрат, содержащий белки, все известные витамины, ценные минеральные вещества, по общему аминокислотному составу обножка близка к другим богатым белком пищевым продуктам — мясу, молоку, яйцам.класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
сборное название насекомых подотряда жалящих перепончатокрылых (кроме пчел и муравьев). Длина до 40 мм (шершни). Ок. 23 тыс. видов. Распространены широко. Опылители растений. Личинки некоторых ос паразиты других насекомых.
необратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.






