






Все без исключения виды рода Formica строят гнезда в почве. Плотность этих гнезд может быть довольно высокой, правда, не такой, как у ![]() Lasius niger или
Lasius niger или ![]() Tetramorium caespitum. Однако в отличие от этих видов глубина, на которую Formica роют ходы, значительно больше. Несомненно, что своей деятельностью они должны оказывать серьезное влияние на почву.
Tetramorium caespitum. Однако в отличие от этих видов глубина, на которую Formica роют ходы, значительно больше. Несомненно, что своей деятельностью они должны оказывать серьезное влияние на почву.
В то же время вопрос о почвообразующей деятельности муравьев вообще и Formica, в частности, изучен пока очень слабо. Имеются лишь отрывочные данные в статьях разных авторов.
Прежде всего остановимся на фактах, свидетельствующих о том, что Formica изменяют почву. Хорошо известно и было описано еще Форелем (Forel, 1874), что растительность вокруг холмиков ![]() F. rufa s. l. резко отличается от окружающей. В сложном ельнике на суглинистых почвах в районе станции Хлюпино (Московская область) мы даже использовали эту особенность для того, чтобы картировать гнезда F. aquilonia. Вокруг холмиков этих муравьев были густые заросли звездчатки (Stellaria sp.), заметные издали. На остальной территории это растение встречалось единично. В. И. Гримальский (1960) пишет, что вокруг гнезд
F. rufa s. l. резко отличается от окружающей. В сложном ельнике на суглинистых почвах в районе станции Хлюпино (Московская область) мы даже использовали эту особенность для того, чтобы картировать гнезда F. aquilonia. Вокруг холмиков этих муравьев были густые заросли звездчатки (Stellaria sp.), заметные издали. На остальной территории это растение встречалось единично. В. И. Гримальский (1960) пишет, что вокруг гнезд ![]() F. rufa (s. 1.) на левобережном Полесье Украины были особо густые заросли орляка, а местами нитрофилов: чистотела (Chelidonium majus) и крапивы (Urtica dioica). Сходные скопления нитрофилов мы также наблюдали в некоторых участках Воронежского заповедника. Невский (по Лебедеву, 1949) наблюдал вокруг гнезд муравейников пышное разрастание костяники (Rubus saxatilis). К. А. Кострин (1951) сообщает, что многие муравейники
F. rufa (s. 1.) на левобережном Полесье Украины были особо густые заросли орляка, а местами нитрофилов: чистотела (Chelidonium majus) и крапивы (Urtica dioica). Сходные скопления нитрофилов мы также наблюдали в некоторых участках Воронежского заповедника. Невский (по Лебедеву, 1949) наблюдал вокруг гнезд муравейников пышное разрастание костяники (Rubus saxatilis). К. А. Кострин (1951) сообщает, что многие муравейники ![]() F. rufa s. 1. окружены зарослями шиповника и малины. Эти растения были вдвое больше тех, которые находились в отдалении от муравейников, и дали больший урожай. Ягоды на этих растениях были сочнее, слаще и в 1,5— 2 раза крупнее.
F. rufa s. 1. окружены зарослями шиповника и малины. Эти растения были вдвое больше тех, которые находились в отдалении от муравейников, и дали больший урожай. Ягоды на этих растениях были сочнее, слаще и в 1,5— 2 раза крупнее.
По наблюдениям А. А. Лебедева (1949), в районе Калинина, Бологого и Вышнего Волочка в почве в основании муравейников рыжих лесных муравьев наблюдается густое сплетение корней ели и березы. Состояние этих деревьев гораздо лучше, чем удаленных от муравейников.
Почвообразующая деятельность муравьев складывается из следующих факторов: перемешивание почвы, изменение механического состава, изменение химического состава почвы. В отношении первых двух сторон деятельности муравьев известно очень мало. Знаем лишь, что муравьи роют ходы и поэтому перемешивают почву и улучшают своими ходами доступ воздуха к корням (Нефедов, 1930; Атанасов 1952). Н. А. Димо (1955) пытался сделать пересчет количества земли, выбрасываемой муравьями, путем взвешивания выбросов из гнезд на глинистых площадках, образовавшихся после половодья. Однако эти данные свидетельствуют лишь о деятельности муравьев по восстановлению разрушенных гнезд, а не о нормальной их деятельности. Воронов (1950) сделал попытку определить почвообразующую деятельность ![]() Lasius niger,
Lasius niger, ![]() L. flavus и
L. flavus и ![]() F. fusca, измеряя гнездовые холмики этих видов. Однако Понтин (Pontin, 1953) наблюдал, как семья
F. fusca, измеряя гнездовые холмики этих видов. Однако Понтин (Pontin, 1953) наблюдал, как семья ![]() L. flavus в течение 5 лет закопала цементную плитку 23x23x1,25 см, но при этом форма и размеры холмика почти не изменились.
L. flavus в течение 5 лет закопала цементную плитку 23x23x1,25 см, но при этом форма и размеры холмика почти не изменились.
ТАБЛИЦА 7
Влияние муравьев на рН кислых подзолистых лесных почв
(Гринфельд, 1941)
|
Вид муравьев
|
рН гнезд
|
рН почвы па глубине (в см)
|
||
|
на поверхности
|
в центре гнезда
|
0—10
|
40—50
|
|
|
6,85
|
6,85
|
6,60
|
6,80
|
|
|
»
|
6,85
|
6,80
|
6,60
|
6,60
|
|
6,85
|
—
|
6,80
|
6,85
|
|
|
»
|
7,00
|
—
|
6,60
|
6,80
|
|
7,00
|
6,95
|
6,60
|
6,80
|
|
|
»
|
6,70
|
6,80
|
6,60
|
6,80
|
|
»
|
6,90
|
7,20
|
6,80
|
6,80
|
|
»
|
6,90
|
—
|
6,80
|
6,85
|
|
7,00
|
7,00
|
6,80
|
6,85
|
|
|
»
|
6,80
|
6,80
|
6,70
|
6,65
|
|
7,00
|
7,55
|
6,90
|
6,85
|
|
|
»
|
6,75
|
7,60
|
6,50
|
6,80
|
Влияние муравьев на рН почвы установлено лишь в самой общей форме. Э. К Гринфельд (1941) обнаружил, что на кислых подзолистых лесных почвах реакция почвы в гнездах ![]() F. rufibarbis (= cunicularia ?),
F. rufibarbis (= cunicularia ?), ![]() F. rufa,
F. rufa, ![]() L. niger,
L. niger, ![]() L. flavus и
L. flavus и ![]() L. umbratus была более щелочной, чем реакция почвы вдали от муравейников (табл.7). В верхних горизонтах рН в гнездах приблизительно на 0,2 выше, т. е. концентрация водородных ионов в гнездах вдвое меньше, чем в почве. Н. А. Крупеников (1951) показал, что на кислых почвах Lasius sp. и Myrmica sp. действительно оказывают такой же эффект, но на щелочных лугово-солончаковых почвах, наоборот, рН гнезд этих муравьев несколько ниже, чем в почве, но изменения в этом случае меньше (табл.8). Аналогичные данные были получены Пиклсом (Pickles, 1941) для
L. umbratus была более щелочной, чем реакция почвы вдали от муравейников (табл.7). В верхних горизонтах рН в гнездах приблизительно на 0,2 выше, т. е. концентрация водородных ионов в гнездах вдвое меньше, чем в почве. Н. А. Крупеников (1951) показал, что на кислых почвах Lasius sp. и Myrmica sp. действительно оказывают такой же эффект, но на щелочных лугово-солончаковых почвах, наоборот, рН гнезд этих муравьев несколько ниже, чем в почве, но изменения в этом случае меньше (табл.8). Аналогичные данные были получены Пиклсом (Pickles, 1941) для ![]() L. flavus.
L. flavus.
О более существенных изменениях муравьями химического состава почвы данных почти нет. В. И. Гримальский показал, что гумусный горизонт с мелкокомковатой структурой под муравейником ![]() F. rufa s. l. достигал толщины 16—17 см, а вне муравейника — всего 5 см. Элювиальный горизонт (13—56 см) под муравейником изобиловал гумусными пятнами, а местами попадались разорванные прослойки гумуса. Вне гнезда гумусные пятна встречались гораздо реже, а гумусных прослоек не обнаружено. Химический анализ показал, что непосредственно у края муравейника содержалось 13,6 мг легко гидролизуемого азота, а в 3 м от гнезда — всего 9,8 мг (Гримальский, 1960).
F. rufa s. l. достигал толщины 16—17 см, а вне муравейника — всего 5 см. Элювиальный горизонт (13—56 см) под муравейником изобиловал гумусными пятнами, а местами попадались разорванные прослойки гумуса. Вне гнезда гумусные пятна встречались гораздо реже, а гумусных прослоек не обнаружено. Химический анализ показал, что непосредственно у края муравейника содержалось 13,6 мг легко гидролизуемого азота, а в 3 м от гнезда — всего 9,8 мг (Гримальский, 1960).
ТАБЛИЦА 8
Влияние муравьев на рН щелочных лугово-солончаковых почв
(Крупеников, 1951)
|
Вид муравьев
|
рН гнезд
|
рН почвы па глубине (в см)
|
||
|
у поверхности
|
в центре гнезда
|
0—10
|
30—40
|
|
| Myrmica sp. |
7,8
|
7,8
|
7,9
|
8,0
|
|
»
|
7,9
|
7,9
|
8,0
|
8,0
|
| Lasius sp. |
7,9
|
8,0
|
8,0
|
8,0
|
|
»
|
7,9
|
7,9
|
7,9
|
8,0
|
При раскопке небольшого гнезда ![]() F. truncorum в Воронежском заповеднике на песчаной почве мы также наблюдали утолщение гумусного слоя под муравейником (рис:69). Но при раскопке гнезд
F. truncorum в Воронежском заповеднике на песчаной почве мы также наблюдали утолщение гумусного слоя под муравейником (рис:69). Но при раскопке гнезд ![]() F. cunicularia glauca, F. cinerea,
F. cunicularia glauca, F. cinerea, ![]() F. sanguinea и
F. sanguinea и ![]() F. pressilabris этого мы не обнаружили. На песчаных почвах вокруг постоянно используемых ходов (на глубине до 70 см) у
F. pressilabris этого мы не обнаружили. На песчаных почвах вокруг постоянно используемых ходов (на глубине до 70 см) у ![]() F. sanguinea и
F. sanguinea и ![]() F. cunicularia glauca на срезе явственно видны кольца гумуса диаметром около 3 см. Возможно, накопление гумуса отчасти объясняется тем, что
F. cunicularia glauca на срезе явственно видны кольца гумуса диаметром около 3 см. Возможно, накопление гумуса отчасти объясняется тем, что ![]() F. cunicularia (а также Cataglyphis aenescens) часто не выбрасывают отбросов пищи из гнезда, а складывают их в специальные камеры (рис:66). Как правило, это овальные камеры длиной до 10 см и высотой 3—5 см, расположенные горизонтально. В одном случае мы наблюдали, как муравьи использовали для этой цели вертикально идущие ходы в нижней части гнезда (рис:66/1/). В том же гнезде в одной камере, с того края, куда открывался вход, помещались личинки, а дальняя ее часть была заполнена пищевыми отбросами. После того, как камера заполняется, муравьи перестают ее посещать. В заброшенных камерах заметно густое переплетение корней растений. Другие виды рода, например все виды Formica s. str. и F. exsecta, по нашим наблюдениям, таких камер не делают, а уносят отбросы на границы территории. F. cinerea выбрасывает остатки хитина добычи, мертвых муравьев и шкурки коконов вместе с землей поблизости от входов.
F. cunicularia (а также Cataglyphis aenescens) часто не выбрасывают отбросов пищи из гнезда, а складывают их в специальные камеры (рис:66). Как правило, это овальные камеры длиной до 10 см и высотой 3—5 см, расположенные горизонтально. В одном случае мы наблюдали, как муравьи использовали для этой цели вертикально идущие ходы в нижней части гнезда (рис:66/1/). В том же гнезде в одной камере, с того края, куда открывался вход, помещались личинки, а дальняя ее часть была заполнена пищевыми отбросами. После того, как камера заполняется, муравьи перестают ее посещать. В заброшенных камерах заметно густое переплетение корней растений. Другие виды рода, например все виды Formica s. str. и F. exsecta, по нашим наблюдениям, таких камер не делают, а уносят отбросы на границы территории. F. cinerea выбрасывает остатки хитина добычи, мертвых муравьев и шкурки коконов вместе с землей поблизости от входов.
Большое значение для выяснения почвообразующей деятельности муравьев имеют данные о том, насколько часто муравьи меняют место гнезда. Относительно Formica имеется только одна работа Шербы (Scherba, 1958), посвященная F. ulkei, однако эти данные мы не можем использовать, так как в Палеарктике не встречается ни одного близкого к F. ulkei вида.
Для выяснения этого вопроса мы провели несколько наблюдений. В 1956 г. в 519-м квартале Воронежского заповедника (сосновая монокультура 30 лет, на старопахоте) на площади около 40 га мы провели полный учет гнезд Formica s. str. и закартировали их. В 1961 г., по просьбе автора, там же провел учет Б. А. Смирнов. В 1963 г. мы снова проделали ту же работу. Доминирующим видом в этом участке был луговой муравей (F. pratensis). В 1956 г. здесь было 32 гнезда этого муравья, в 1961 г.— 20 и в 1963 г.— 22. С 1956 г. на их прежнем месте осталось только 4 гнезда (12,5%), а с 1961 — 15 гнезд (75%). Таким образом, ежегодно по различным причинам (перекочевывали или погибали) покидалось 10—20% муравейников.
В 1962 и 1963 гг. мы провели картирование 3 колоний и нескольких отдельных гнезд ![]() F. rufa и F. polyctena в смешанном лесу на суглинках в районе станции Жаворонки Московской области. В 1962 г. здесь было 19 гнезд, а в 1963 г. — 20. Но не изменилось положение только у 14 гнезд (75%). Проведя на основании этих данных элементарные подсчеты, можно прийти к выводу, что роль Formica, особенно рыжих лесных муравьев, в механическом перемешивании почвы невелика. Однако гнезда Formica в лесах могут служить очагами расселения ряда растений. Например, в Воронежском заповеднике мы наблюдали, что возобновление дуба в суборях идет главным образом по периферии муравейников рыжих лесных муравьев.
F. rufa и F. polyctena в смешанном лесу на суглинках в районе станции Жаворонки Московской области. В 1962 г. здесь было 19 гнезд, а в 1963 г. — 20. Но не изменилось положение только у 14 гнезд (75%). Проведя на основании этих данных элементарные подсчеты, можно прийти к выводу, что роль Formica, особенно рыжих лесных муравьев, в механическом перемешивании почвы невелика. Однако гнезда Formica в лесах могут служить очагами расселения ряда растений. Например, в Воронежском заповеднике мы наблюдали, что возобновление дуба в суборях идет главным образом по периферии муравейников рыжих лесных муравьев.
Почвообразующая деятельность других муравьев, например некоторых видов из родов Lasius, Myrmica, Tetramorium и других, очевидно, гораздо значительнее, так как плотность их в некоторых местах достигает одного гнезда на 1 м², и старые гнезда покидаются их населением, по-видимому, чаще, чем у Formica (Нефедов, 1930; Pontin, 1963; Oinonen, 1956).
Муравьи являются пионерами обнаженных склонов гор (Атанасов, 1952) и скал (Oinonen, 1956) и играют важную роль в их обрастании. Правда, роль Formica здесь незначительна. Первым из муравьев на скалах в южной Финляндии поселяется ![]() F. rufibarbis (Oinonen, 1. с).
F. rufibarbis (Oinonen, 1. с).
В разложении древесины и превращении ее в почву муравьи играют важную роль. Б. М. Мамаев (1960) выделяет следующие стадии разрушения древесины: 1) церамбицидная, 2) люканидная, 3) формицидная и 4) люмбрицидная. На третьей стадии основную роль играют муравьи, главным образом виды из родов Lasius, Camponotus. Formica имеют меньшее значение. Из видов этого рода в разрушении древесины принимают участие ![]() F. rufa,
F. rufa, ![]() F. truncorum,
F. truncorum, ![]() F. fusca (в конце формицидной стадии),
F. fusca (в конце формицидной стадии), ![]() F. sanguinea (в горных лесах Средней Азии), реже F. aquilonia, F. lugubris и F. polyctena.
F. sanguinea (в горных лесах Средней Азии), реже F. aquilonia, F. lugubris и F. polyctena.
Муравьи довольно строго приурочены к типу и влажности почвы (Атанасов, 1952; Бальц, 1915; Нефедов, 1930) и могут служить индикатором почвенных условий. Однако (во всяком случае для Formica) это вряд ли может найти практическое применение, так как растения являются лучшими индикаторами, а главное определить их гораздо легче, чем Formica.

Рис. 69. Строение небольшого гнезда
1 — гумусный слой почвы; 2 — древесина пня; 3 — камеры с рабочими и личинками; 4 — камеры с крылатыми половыми особями; 5 — гнездовые камеры Formicoxenus nitidulus; 6 — местонахождения живых личинок Clytra quadripunctata; 7 — местонахождения пустых старых чехликов личинок С. quadripunctata; ♀ — камера с «царицей»; x — насыпной холмик из растительных остатков; I — схема, поясняющая, как брались разрезы гнезда II и III
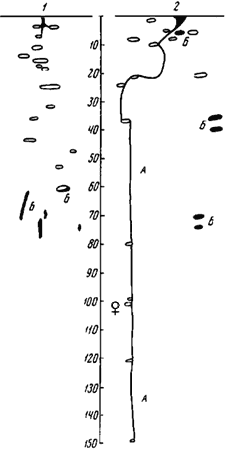
Рис. 66. Строение двух гнезд
А — зимовочный ход; Б — камеры с отбросами; ♀ — камера с самкой. На вертикальной шкале — глубина (в см)
 Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant
Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant Tetramorium caespitum (Linnaeus, 1758) — дерновый муравей, pavement ant
Tetramorium caespitum (Linnaeus, 1758) — дерновый муравей, pavement antСтарое название: T. indocile
 Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравей Lasius flavus (Fabricius, 1782) — желтый луговой (садовый, земляной) муравей
Lasius flavus (Fabricius, 1782) — желтый луговой (садовый, земляной) муравей Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant
Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky antосновная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
 Formica rufibarbis Fabricius, 1793 — краснощекий муравей, краснощекий муравей-минер
Formica rufibarbis Fabricius, 1793 — краснощекий муравей, краснощекий муравей-минер Lasius umbratus (Nylander, 1846) — желтый пахучий муравей, пахучий земляной муравей
Lasius umbratus (Nylander, 1846) — желтый пахучий муравей, пахучий земляной муравейВсего 100 видов.
 Formica truncorum Fabricius, 1804 — красноголовый муравей
Formica truncorum Fabricius, 1804 — красноголовый муравей Formica cunicularia Latreille, 1798 — прыткий степной муравей
Formica cunicularia Latreille, 1798 — прыткий степной муравей Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелец
Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелец Formica pressilabris Nylander, 1846 — малый тонкоголовый лесной муравей
Formica pressilabris Nylander, 1846 — малый тонкоголовый лесной муравейполисахарид, образованный остатками аминосахара ацетилглюкозамина. Основной компонент наружного скелета (кутикулы) насекомых, ракообразных и других членистоногих. У грибов заменяет целлюлозу, с которой сходен по химическим и физическим свойствам и биологической роли.
функциональное образование, состоящее из материнской и дочерних семей, поддерживающих регулярные родственные отношения
Всего 415 видов
Всего 965 видов.






