






Регуляция температуры летом. Для развития молоди муравьев нужен оптимальный режим влажности и температуры. В табл.4 приводятся данные исследований Гринфельда /Здесь и дальше мы даем современные названия видов, с которыми работал этот автор./ (1939), полученные в опытах с термоградиентами, свидетельствующие о довольно высоком температурном оптимуме большинства Formica. Ниже всех лежит оптимум у ![]() F. fusca. Для сравнения приведены данные о предпочитаемых температурах у типичных лесных видов:
F. fusca. Для сравнения приведены данные о предпочитаемых температурах у типичных лесных видов: ![]() Myrmica rubra, М. scabrinodis,
Myrmica rubra, М. scabrinodis, ![]() Lasius niger. Следует, однако, оговорить, что предпочитаемая температура зависит от температуры, при которой муравьи выращивались. Так, особи
Lasius niger. Следует, однако, оговорить, что предпочитаемая температура зависит от температуры, при которой муравьи выращивались. Так, особи ![]() F. rufa s. l., выращенные при 3— 4°, предпочитают температуру 23—24°, а выращенные при 25—27° — температуру 31—32° (Herter, 1924).
F. rufa s. l., выращенные при 3— 4°, предпочитают температуру 23—24°, а выращенные при 25—27° — температуру 31—32° (Herter, 1924).
ТАБЛИЦА 4
Температуры, предпочитаемые муравьями для выращивания молоди
(по Гринфельду, 1939)
|
Виды муравьев
|
Число опытов
|
Температура °C
|
|||
|
min
|
max
|
max для кукол
|
оптимум
|
||
|
20
|
24
|
32
|
36
|
30—32
|
|
| F. cinerea imitans |
18
|
25
|
31
|
34
|
29—30
|
|
15
|
22—23
|
29—30
|
35
|
27—28
|
|
|
11
|
23
|
34
|
—
|
29—32
|
|
| F. exsecta |
10
|
—
|
30
|
—
|
29—30
|
|
16
|
18
|
24
|
—
|
23—24
|
|
| M. scabrinodis |
13
|
21
|
26
|
—
|
24—25
|
|
22
|
25
|
28
|
—
|
27—28
|
|
Земляные гнезда муравьев можно разделить по их температурному режиму на две группы: 1) гнезда, температура которых не отличается от температуры окружающей среды, и 2) гнезда, температура которых выше температуры окружающей среды. К первой группе относятся земляные гнезда без наружных построек. В этих гнездах муравьи находят оптимальные условия для молоди, перенося ее в разные слои почвы и поселяясь в таких стациях, где температурный режим почвы больше всего подходит для них (Гринфельд, 1939; Steiner, 1947).
ТАБЛИЦА 5
Средняя температура 10 холмиков ![]() Formica cunicularia glauca (средняя высота 16 см) и почвы рядом с гнездами. Заповедник «Лес на Ворскле»
Formica cunicularia glauca (средняя высота 16 см) и почвы рядом с гнездами. Заповедник «Лес на Ворскле»
(Гринфельд, 1939)
|
Время, часы
|
Температура °C
|
||||||||||
|
воздуха
|
в холмике на глубине (в см)
|
почвы на глубине (в см)
|
|||||||||
|
0
|
3—5
|
10
|
20
|
30
|
0
|
3—5
|
10
|
20
|
30
|
||
|
7
|
18,4
|
18,0
|
18,4
|
18,5
|
19,0
|
19,7
|
18,3
|
19,4
|
19,5
|
19,8
|
19,8
|
|
13
|
28,0
|
38,5
|
30,1
|
26,6
|
22,3
|
20,5
|
27,5
|
25,6
|
20,2
|
20,5
|
20,1
|
|
21
|
19,3
|
19,5
|
21,5
|
23,8
|
23,5
|
21,5
|
21,5
|
22,2
|
21,5
|
21,6
|
20,7
|
Ко второй группе относятся гнезда с примитивными земляными холмиками. Благодаря тому, что холмики возвышаются под травой, почва в них суше (меньше теплопроводность почвы) и наклонные скаты холмика лучше прогреваются солнцем, поэтому температура холмиков, хотя и варьирует, но обычно выше, чем температура почвы. В табл.5 приведены данные о средней температуре 10 холмиков (средняя высота 16 см) ![]() F. cunicularia glauca на разной глубине и температура почвы рядом с гнездами на такой же глубине (Гринфельд, 1939). Из приведенных данных видно, что на глубине 0—5 см в холмике в середине дня температура была выше оптимальной, тогда как в почве она за весь день не превысила 30°. На глубине свыше 30 см разницы уже практически нет. Сходные данные были получены для холмиков североамериканских F. exsectoides и F. ulkei (Scherba, 1962; Williams, 1961).
F. cunicularia glauca на разной глубине и температура почвы рядом с гнездами на такой же глубине (Гринфельд, 1939). Из приведенных данных видно, что на глубине 0—5 см в холмике в середине дня температура была выше оптимальной, тогда как в почве она за весь день не превысила 30°. На глубине свыше 30 см разницы уже практически нет. Сходные данные были получены для холмиков североамериканских F. exsectoides и F. ulkei (Scherba, 1962; Williams, 1961).
Таким образом, гнезда с холмиками позволяют муравьям поддерживать в них больший градиент температуры. Кроме того, как уже говорилось выше, муравьи в зависимости от условий могут строить разные гнезда. Так, по данным Гринфельда (1939), в заповеднике «Лес на Ворскле» на безлесном южном склоне (за лето максимальная температура почвы в 13 час. на глубине 10 см была 35°) ![]() F. cunicularia glauca строили гнезда исключительно без холмиков, а на полянах в дубовом лесу (максимальная температура на той же глубине 22°) из 12 найденных гнезд 10 были с холмиками.
F. cunicularia glauca строили гнезда исключительно без холмиков, а на полянах в дубовом лесу (максимальная температура на той же глубине 22°) из 12 найденных гнезд 10 были с холмиками.
Уже давно было замечено, что в холодную погоду температура в гнездовых холмиках рыжих лесных муравьев выше температуры воздуха и почвы (Forel, 1874; Wheeler, 1910). Разница между температурой воздуха и температурой внутри гнездовых холмиков может достигать более 20° (Гринфельд, 1939). Но характерной особенностью этих гнезд является относительное постоянство режима температуры в течение всего периода выведения молоди. В холмиках F. polyctena, например, температура все лето поддерживается в среднем между 23 и 30° (максимальные отклонения 18—33°). Наименее изменчива температура на глубине около 30 см (Raigner, 1947, 1948). У ![]() F. rufa rufopratensis (rufa ?) регуляция температуры наблюдается в зоне от 15 до 50 см под поверхностью купола, и здесь градиент температур от 23 до 29°. Наиболее постоянной (около 26°) сохраняется температура на глубине 30 см (как и у F. polyctena). Средняя температура почвы на той же глубине была 16° (Steiner, 1924, 1947). Сходные данные были получены Гринфельдом (1939) для
F. rufa rufopratensis (rufa ?) регуляция температуры наблюдается в зоне от 15 до 50 см под поверхностью купола, и здесь градиент температур от 23 до 29°. Наиболее постоянной (около 26°) сохраняется температура на глубине 30 см (как и у F. polyctena). Средняя температура почвы на той же глубине была 16° (Steiner, 1924, 1947). Сходные данные были получены Гринфельдом (1939) для ![]() F. rufa, но в связи с тем, что он брал небольшие гнезда (средняя высота 30 см), регуляция наблюдалась в зоне 10—30 см, и градиент температур был меньше (24,7—29,4°). Аналогичные данные получил Като (Kato, 1939) для F. lugubris (=
F. rufa, но в связи с тем, что он брал небольшие гнезда (средняя высота 30 см), регуляция наблюдалась в зоне 10—30 см, и градиент температур был меньше (24,7—29,4°). Аналогичные данные получил Като (Kato, 1939) для F. lugubris (=![]() F. truncorum yessensis) в Японии.
F. truncorum yessensis) в Японии.
Первые исследователи считали, что такая разница возникает главным образом в результате химического нагрева гнезда при разложении растительных остатков, из которых состоит купол (Forel, 1874; Wheeler, 1910). Сейчас это мнение опровергнуто, так как доказано, что при отсутствии муравьев температурный режим резко отличается от режима населенных гнезд и приближается к режиму земляных холмиков: температура в брошенных муравейниках ниже, чем в жилых (Steiner, 1947; Raigner, 1947). Э. К. Гринфельд (1939) провел следующий опыт: выбрав два одинаковых гнезда ![]() F. rufa, он посыпал одно из гнезд нафталином, и муравьи его покинули. Через несколько дней в 13 час. была измерена температура в обоих гнездах. Она была на глубине 10 см в населенном гнезде 28,4° и в брошенном — 22,3°, на глубине 20 см, соответственно, 28,0 и 19,5°, на глубине 30 см — 27,0 и 18,0° и на глубине 40 см — 24,6 и 18,0°. В это время температура воздуха была 28,7°, а температура почвы на глубине 10 см — 18,1°.
F. rufa, он посыпал одно из гнезд нафталином, и муравьи его покинули. Через несколько дней в 13 час. была измерена температура в обоих гнездах. Она была на глубине 10 см в населенном гнезде 28,4° и в брошенном — 22,3°, на глубине 20 см, соответственно, 28,0 и 19,5°, на глубине 30 см — 27,0 и 18,0° и на глубине 40 см — 24,6 и 18,0°. В это время температура воздуха была 28,7°, а температура почвы на глубине 10 см — 18,1°.
Наиболее важным фактором терморегуляции у рыжих лесных муравьев является строение гнездового купола. Форма купола каждого муравейника оптимальна для данных условий, и если искусственно изменить ее, перенеся часть материала из одних участков в другие, муравьи быстро восстанавливают прежнюю форму (Chauvin, 1960). Ланге (Lange, 1959а) поставил следующий эксперимент: три искусственных гнезда F. polyctena нагревали сверху при помощи инфракрасных ламп мощностью 150 вт, установленных на высоте 48 см от основания гнезда. Через 30 дней форма холмика стабилизировалась. Купол оказался сильно уплощенным. На 31-й день опыта лампы были подняты на высоту 100 см; 10 дней спустя муравьи во всех формикариях выстроили высокие купола. Через 41 день после начала опыта лампы были опущены до 36 см и после этого форма купола вновь была изменена на плоскую.
Полевые наблюдения также подтверждают эту закономерность. Уже давно было подмечено (Forel, 1874; Wheeler, 1910), что холмики Formica имеют, как правило, восточную и южную экспозицию (чтобы прогрев купола обеспечил раннее начало активности), и в затененных местах они выше, чем на открытых. Но, как справедливо заметил Экштайн (Eckstein, 1937), из этого правила часто встречаются исключения. Однако они вызываются другими причинами, такими, как средняя температура почвы или плотность населения в гнезде, но общая закономерность остается.
Шерба (Scherba, 1958), изучавший корреляцию между направлением длинного ската гнезда и основным направлением солнечного освещения у F. ulkei в районе Чикаго, статистически доказал, что гнезда, находящиеся на открытых местах или затененные с северо-востока, востока и юго-востока, ориентированы приблизительно на юго-запад, а затененные с юго-запада, запада и северо-запада — приблизительно на юго-восток.
Летом 1962 г. в Воронежском заповеднике мы провели серию измерений, чтобы обнаружить зависимость между наклоном длинного (обычно южного) ската куполов ![]() F. rufa, F. exsecta и F. pratensis и количеством прямого солнечного света, падающего на гнездо.
F. rufa, F. exsecta и F. pratensis и количеством прямого солнечного света, падающего на гнездо.
Определение количества солнечного света проводилось следующим образом. При помощи астрономической таблицы положения солнца (на 25 июня для широты 51°), компаса и угломера мысленно проводилась траектория солнца, видимая с поверхности муравейника, и отмечались часы, когда солнце освещает гнездо (т. е. не загорожено ветвями деревьев и т. п.). Затем, исходя из данных об интенсивности солнечной радиации в течение суток, была составлена таблица, по которой определялось, какое относительное количество солнечной радиации получает горизонтальная поверхность в разные часы дня (на 21 июня). Сопоставляя полученные данные с этой таблицей, можно было с точностью до 10% определить, какое количество прямых солнечных лучей получает гнездо за сутки, если принять максимальное возможное количество за 100%. Наклон длинного ската определялся по угломеру геологического компаса.
Таким методом было исследовано 60 гнезд F. pratensis, 50 гнезд ![]() F. rufa и 30 гнезд F. exsecta.
F. rufa и 30 гнезд F. exsecta.
Результаты исследований изображены на графиках (рис:41, рис:70). Из полученных данных видно, что, несмотря на большую изменчивость, линии регрессии показывают явную корреляцию между крутизной ската и количеством прямого солнечного света: с увеличением освещенности гнезда становятся более плоскими. Это вполне естественно, так как приближение нагреваемого ската к перпендикуляру по отношению к направлению лучей солнца (для Воронежского заповедника — к 52°) обусловливает большее нагревание поверхности гнезда (Starke, 1943) /Штерке высказал предположение о наличии этой связи, но не подтвердил его никакими цифрами./.
Ближе всего эта зависимость выражается формулой
Sin β = a + bR,
где β — угол наклона длинного ската гнезда; R — количество солнечной радиации, падающей на горизонтальную поверхность, находящуюся на куполе, в процентах от максимальной радиации (на открытом месте); а — величина положительная; b — величина отрицательная. В зависимости от экологических особенностей видов (или экологических рас) степень этой зависимости может варьировать: у ![]() F. rufa она выражается уравнением Sin β = 0,69—0,026 R, у F. exsecta — Sin β = 0,56—0,0024 R, у расы nigricans F. pratensis — Sin β = 1,165—0,0119 R, у типичной F. pratensis — Sin β = 0,528—0,0039 R.
F. rufa она выражается уравнением Sin β = 0,69—0,026 R, у F. exsecta — Sin β = 0,56—0,0024 R, у расы nigricans F. pratensis — Sin β = 1,165—0,0119 R, у типичной F. pratensis — Sin β = 0,528—0,0039 R.
Рис. 70. Зависимость угла наклона (β) пологого ската купола гнезд F. rufа (А) и F. exsecta (Б) от количества прямой солнечной радиации (R, %) (Воронежский заповедник)
Как показали исследования Ренье (Raigner, 1947) на F. polyctena, для регуляции температуры большое значение имеет структура наружных слоев купола. В жаркие Дни с поверхности купола происходит усиленное испарение, чему способствует открывание специальных испаряющих камер (которые также служат для вентиляции гнезда). В результате этого температура в гнезде понижается. Ночью в камерах и ходах наружных слоев купола происходит конденсация влаги и образуется туман, создающий дополнительную теплоизолирующую защиту.
Штайнер (Steiner, 1930, 1947) считал, что определенную, хотя и незначительную, роль играет тепло, выделяемое рабочими муравьями. В начале ночи температура в гнезде повышается на 3°. Этот факт автор объяснял тем, что муравьи, собираясь вечером в гнездо, нагревают его. Однако Ренье (Raigner, 1947) связывал этот факт с образованием теплоизолирующего слоя при понижении температуры воздуха, в результате чего происходит нагревание верхних слоев под влиянием еще теплой внутренней части гнезда.
По наблюдениям Э. К. Гринфельда (1939) и других авторов, температура гнезд ![]() F. rufa с большим населением выше, чем температура малонаселенных гнезд (рис:71). Штайнер (Steiner, 1929) обнаружил, что в холмиках F. exsecta с большим количеством молоди температура бывает выше. Эти авторы считают, что такие различия обусловливаются непосредственно физиологическим теплом муравьев. Однако возможен и другой вариант: большее количество муравьев способно лучше регулировать режим гнезда, быстро перестраивая его соответственно обстановке.
F. rufa с большим населением выше, чем температура малонаселенных гнезд (рис:71). Штайнер (Steiner, 1929) обнаружил, что в холмиках F. exsecta с большим количеством молоди температура бывает выше. Эти авторы считают, что такие различия обусловливаются непосредственно физиологическим теплом муравьев. Однако возможен и другой вариант: большее количество муравьев способно лучше регулировать режим гнезда, быстро перестраивая его соответственно обстановке.
Таким образом, вопрос о роли физиологического тепла, выделяемого самими муравьями, остается открытым.
Регуляция влажности. Э. К. Гринфельд (1939) при выяснении оптимального режима влажности для выведения молоди у муравьев поставил серию опытов, используя прибор, в котором создавался градиент влажности. Из данных опытов (табл.6) видно, что Formica, как и все другие муравьи, выбирают, как правило, наибольшую влажность почвы. Эти данные согласуются с результатами исследований Гессвальда (Gosswald, 1938b), показавшего, что для развития молоди муравьев нужна 100%-ная относительная влажность воздуха /Вообще почти для всех почвенных животных оптимальная влажность воздуха порядка 100% (Гиляров, 1949)./.
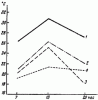
Рис. 71. Регуляция температуры в холмиках F. rиfа с различной плотностью населения (по Гринфельду, 1939)
1 — максимальная температура густонаселенного (старого) гнезда; 2 — то же молодого гнезда; 3 — температура воздуха; 4 — температура почвы на глубине 10 см
В опытах с градиентом температуры (Herter, 1924; Гринфельд, 1939) влажность почвы роли не играет, хотя в опытах Гринфельда она менялась от 2 до 27% (абсолютная влажность почвы). Известно также, что некоторые виды, изученные Гринфельдом (F. cinerea, ![]() F. cunicularia glauca), обитают в условиях Курской области (заповедник «Лес па Ворскле») в наиболее ксерофитных стациях. Исходя из этого, Гринфельд делает вывод, что муравьи являются в принципе абсолютными гигрофилами, но требования к высокой температуре для развития молоди заставляют их селиться в сухих местах (т. е. температура является минимум-фактором в их распределении).
F. cunicularia glauca), обитают в условиях Курской области (заповедник «Лес па Ворскле») в наиболее ксерофитных стациях. Исходя из этого, Гринфельд делает вывод, что муравьи являются в принципе абсолютными гигрофилами, но требования к высокой температуре для развития молоди заставляют их селиться в сухих местах (т. е. температура является минимум-фактором в их распределении).
ТАБЛИЦА 6
Предпочитаемая влажность почвы у муравьев рода Formica
(по Гринфельду, 1939)
|
Вид муравьев
|
Максимальная влагоемкость, %
|
Выбрана почва
с влагоемкостью, % |
Расплод
|
|
|
почвы
|
при опыте
|
|||
|
25,0
|
11,1
|
11,1
|
Куколки
|
|
|
»
|
53,0
|
25,0
|
25,0
|
»
|
| F. сinerea imitans |
25,0
|
15,4
|
15,4
|
|
|
»
|
25,0
|
19,2
|
19,2
|
|
|
25,0
|
15,4
|
15,4
|
Личинки и куколки
|
|
|
»
|
25,0
|
15,4
|
15,4
|
То же
|
|
25,0
|
23,1
|
19,2
|
Куколки
|
|
| F. exsecta |
53,0
|
30,0
|
25,0
|
»
|
В гнездах рыжих лесных муравьев (![]() F. rufa) относительная влажность варьирует от 30 до 100%, независимо от температуры (Raigner, 1947).
F. rufa) относительная влажность варьирует от 30 до 100%, независимо от температуры (Raigner, 1947).
У североамериканской F. ulkei, как показали исследования Шербы (Scherba, 1959), напротив, поддерживается постоянный режим влажности. На глубине 5 см под поверхностью купола влажность была в среднем 27,23±0,66%. а на глубине 30 см — 29,40±0,54%. В течение 10 месяцев активности муравьев сезонные колебания составили всего 5,4±1,13%. В то же время в почве рядом с гнездом влажность колебалась от 15,3 до 40,5% на глубине 5 см и от 21,6 до 40,7% на глубине 30 см. Опыты в приборе с градиентом влажности показали, что предпочитаемая влажность для личинок этого муравья 29,58±0,04%, а для куколок — 27,58±0,04%, что почти совпадает с влажностью в холмиках. Механизмы этой регуляции до сих пор совершенно неясны.
Интересны гнезда, которые строит F. picea на торфяных болотах в Европе (Bonner, 1914, 1915; Skwarra, 1929а, b). В этих условиях высокой влажности F. picea строит своеобразные холмики из стеблей мха, поднятые над поверхностью почвы. В тех же условиях верховых болот другие виды делают гнезда в высоких кочках (Маавара, 1955; Рорр, 1962).
Некоторые виды муравьев способны переносить затопление. Так ![]() F. fusca,
F. fusca, ![]() F. cunicularia и
F. cunicularia и ![]() F. sanguinea часто встречаются на заливных лугах и затопляемых весной долинах. Выживанию муравьев в эти периоды способствует то, что в камерах остаются пузырьки воздуха, а так как активность дыхательных процессов в холодной почве у муравьев низка, им этого воздуха бывает достаточно. Даже в активном состоянии муравьи Lasius и Camponotus выживают при погружении их под воду на срок до 3 суток (Fielde, 1904). Однако если муравьев перед погружением обтереть и удалить пузырьки воздуха, застревающие между волосками, они очень быстро погибают (Devaux, по Кузнецову, 1953).
F. sanguinea часто встречаются на заливных лугах и затопляемых весной долинах. Выживанию муравьев в эти периоды способствует то, что в камерах остаются пузырьки воздуха, а так как активность дыхательных процессов в холодной почве у муравьев низка, им этого воздуха бывает достаточно. Даже в активном состоянии муравьи Lasius и Camponotus выживают при погружении их под воду на срок до 3 суток (Fielde, 1904). Однако если муравьев перед погружением обтереть и удалить пузырьки воздуха, застревающие между волосками, они очень быстро погибают (Devaux, по Кузнецову, 1953).
Зимовка. У всех видов Formica зимуют только взрослые особи.
Осенью рыжие лесные муравьи уходят в глубь почвы, где температура не падает ниже +5° (Zahn, 1958). У ![]() F. rufa большая часть особей находится в нижней половине древесного пня, являющегося основой гнезда (Gosswald, 1940d). У
F. rufa большая часть особей находится в нижней половине древесного пня, являющегося основой гнезда (Gosswald, 1940d). У ![]() F. cunicularia и
F. cunicularia и ![]() F. pressilabris, как мы наблюдали в Воронежском заповеднике, рабочие вместе с самкой собираются очень тесными группами в камерах зимовочного хода. Однажды я обнаружил в Московской области зимующее гнездо
F. pressilabris, как мы наблюдали в Воронежском заповеднике, рабочие вместе с самкой собираются очень тесными группами в камерах зимовочного хода. Однажды я обнаружил в Московской области зимующее гнездо ![]() F. fusca. Почти все особи собрались в одной большой камере, образовав некоторое подобие пчелиного клуба. В центре этого клуба находилась «царица» и несколько бескрылых тлей.
F. fusca. Почти все особи собрались в одной большой камере, образовав некоторое подобие пчелиного клуба. В центре этого клуба находилась «царица» и несколько бескрылых тлей.
Зимой муравьи находятся в состоянии диапаузы. Интенсивность жизненных процессов у них резко падает. У F. ulkei рабочие зимой потребляют в 20 раз меньше кислорода, чем рабочие, находящиеся летом при 0° (Dreyer, 1932).
На зимовку рабочие ![]() F. rufa уходят с наполненным зобом и большим количеством жира (Gosswald, 1940а). Сухой вес
F. rufa уходят с наполненным зобом и большим количеством жира (Gosswald, 1940а). Сухой вес ![]() F. rufa, F. polyctena, F. pratensis и F. exsecta в течение зимы (с октября по март) несколько увеличивается по сравнению с общим весом. Это связано с тем, что жидкое содержимое зоба расходуется, а жировое тело остается нетронутым и является резервом для весеннего выкармливания половых особей (Kirchner, 1964).
F. rufa, F. polyctena, F. pratensis и F. exsecta в течение зимы (с октября по март) несколько увеличивается по сравнению с общим весом. Это связано с тем, что жидкое содержимое зоба расходуется, а жировое тело остается нетронутым и является резервом для весеннего выкармливания половых особей (Kirchner, 1964).
Весенний режим температуры в гнездах рыжих лесных муравьев. Ранней весной, как только стает снег, на поверхности гнезд рыжих лесных муравьев появляется большое количество рабочих. Они толстым слоем покрывают освещенные склоны холмиков. Нагревшиеся на солнце особи через 10—20 мин. уходят в гнездо и там, остывая, отдают полученное тепло. Разница в температуре тела рабочих, поднимающихся на поверхность и опускающихся в гнездо, составляет 10—15°. Эти особи получили название «теплоносцев» («Warmetragern»). В результате их деятельности под поверхностью купола образуется «тепловое ядро» с температурой 26—29° (Zahn, 1958). Такой ранневесенний нагрев части гнезда позволяет рыжим лесным муравьям к концу весны вырастить половых особей.
Как показали исследования Гессвальда и Вира (Gosswald, Bier, 1954b), в формикарии, где был создан градиент температуры, самки F. polyctena после пробуждения из зимней спячки поднимаются в теплую часть гнезда. Здесь они откладывают свои «зимние» яйца, из которых затем при нормальных условиях вырастают половые особи. После откладки яиц самки снова уходят в холодную часть гнезда. Такие миграции необходимы. Если содержать муравьев в формикарии с постоянной температурой, из яиц выводятся только рабочие особи.
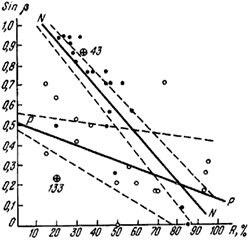
Рис. 41. Зависимость угла наклона пологого ската купола гнезд F. pratensis (β) от количества прямой солнечной радиации (R,%)
N — N, ● — <nigricans>; Р—P, ○ — pratensis; ☺ — гнезда с промежуточными самками. Пунктиром показаны границы изменчивости линии регрессии в пределах ошибки коэффициентов a и b уравнения Sinβ = а + bR

Рис. 70. Зависимость угла наклона (β) пологого ската купола гнезд F. rufа (А) и F. exsecta (Б) от количества прямой солнечной радиации (R, %) (Воронежский заповедник)

Рис. 71. Регуляция температуры в холмиках F. rиfа с различной плотностью населения (по Гринфельду, 1939)
1 — максимальная температура густонаселенного (старого) гнезда; 2 — то же молодого гнезда; 3 — температура воздуха; 4 — температура почвы на глубине 10 см
 Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant
Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant Myrmica rubra (Linnaeus, 1758) — рыжая мирмика
Myrmica rubra (Linnaeus, 1758) — рыжая мирмика Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant
Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравей Formica cunicularia Latreille, 1798 — прыткий степной муравей
Formica cunicularia Latreille, 1798 — прыткий степной муравей Formica truncorum Fabricius, 1804 — красноголовый муравей
Formica truncorum Fabricius, 1804 — красноголовый муравейSubspecies: Myrmica scabrinodis ussuriensis, Kuznetsov-Ugamsky
искусственный муравейник.
(от греч. oikos дом, жилище, местопребывание и ...логия), наука об отношениях живых организмов и образуемых ими сообществ между собой и с окружающей средой. Термин «экология» предложен в 1866 Э. Геккелем. Объектами экологии могут быть популяции организмов, виды, сообщества, экосистемы и биосфера в целом.
организм, предпочитающий влажные местообитания - любящий влажность.
 Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелец
Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелецВсего 100 видов.
Всего 965 видов.
 Formica pressilabris Nylander, 1846 — малый тонкоголовый лесной муравей
Formica pressilabris Nylander, 1846 — малый тонкоголовый лесной муравейнадсемейство летающих насекомых отряда перепончатокрылых, родственное осам и муравьям. Существует около 20 тысяч видов пчёл. Распространены на всех континентах, кроме Антарктиды. Пчёлы питаются нектаром и пыльцой, используя нектар главным образом в качестве источника энергии, а пыльцу для получения белков и других питательных веществ.
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.






