






Еще из опытов Гюбера, Фореля и Леббока (Huber, 1810; Forel, 1874; Леббок, 1898) стало ясно, что муравьи каким-то образом передают друг другу сведения о месте нахождения добычи, а, возможно, также и о ее приблизительном количестве. Например, Леббок часто, описывая поведение ![]() Lasius niger, сообщает: «Муравей вернулся к кормушке и привел своего товарища». Такого рода случаи описывались многократно и впоследствии. Приведу несколько собственных наблюдений.
Lasius niger, сообщает: «Муравей вернулся к кормушке и привел своего товарища». Такого рода случаи описывались многократно и впоследствии. Приведу несколько собственных наблюдений.
Если в искусственных гнездах ![]() L. niger предложить муравьям раствор сахара, он может долго стоять нетронутым. Но стоит одному рабочему найти сахар и, вернувшись в гнездо, покормить других, как спустя полчаса кормушка оказывается полной муравьев. При проведении описанных выше опытов со светокомпасной ориентацией
L. niger предложить муравьям раствор сахара, он может долго стоять нетронутым. Но стоит одному рабочему найти сахар и, вернувшись в гнездо, покормить других, как спустя полчаса кормушка оказывается полной муравьев. При проведении описанных выше опытов со светокомпасной ориентацией ![]() F. sanguinea постоянно приходилось наблюдать, что на арене, которая до этого часами была пустой, за короткое время появлялось около десятка рабочих после того, как один из муравьев (вытащенный специально из входа в гнездо) приносил в гнездо несколько куколок. Любопытно, что даже попав на кормушку с куколками, эти вышедшие из гнезда рабочие куколок не таскают, но в то же время охотно кормятся на кормушке с раствором меда, т.е. в данном случае мобилизация не специфична и из гнезда выходят представители различных полиэтических групп фуражиров. С другой стороны, если постоянно убирать рабочих, перетаскивающих куколки, оставляя только одного, из гнезда все время появлялись все новые и новые рабочие, которые начинали таскать куколок. Следует отметить, что в нашем экспериментальном гнезде
F. sanguinea постоянно приходилось наблюдать, что на арене, которая до этого часами была пустой, за короткое время появлялось около десятка рабочих после того, как один из муравьев (вытащенный специально из входа в гнездо) приносил в гнездо несколько куколок. Любопытно, что даже попав на кормушку с куколками, эти вышедшие из гнезда рабочие куколок не таскают, но в то же время охотно кормятся на кормушке с раствором меда, т.е. в данном случае мобилизация не специфична и из гнезда выходят представители различных полиэтических групп фуражиров. С другой стороны, если постоянно убирать рабочих, перетаскивающих куколки, оставляя только одного, из гнезда все время появлялись все новые и новые рабочие, которые начинали таскать куколок. Следует отметить, что в нашем экспериментальном гнезде ![]() F. sanguinea с 200 рабочими переноской куколок занималось одновременно, как правило, 2 рабочих, во всяком случае не более четырех, хотя мечение показало, что общее число рабочих, относящихся к этой полиэтической группе, было около десятка.
F. sanguinea с 200 рабочими переноской куколок занималось одновременно, как правило, 2 рабочих, во всяком случае не более четырех, хотя мечение показало, что общее число рабочих, относящихся к этой полиэтической группе, было около десятка.
Васман (Wasmann, 1899) первым открыл у муравьев наличие так называемого «антеннального кода». Согласно его теории, муравьи обмениваются информацией, ударяя друг друга усиками и передними ногами. Впоследствии изучению «антеннального кода» был посвящен ряд исследований.
Как показали исследования английского мирмеколога Уоллиса (Wallis, 1961), тактильные стимулы имеют большое значение при трофаллаксисе. Направление обмена пищей у ![]() F. sanguinea и
F. sanguinea и ![]() F. fusca зависит от того, движения какого из партнеров являются более интенсивными. Прикосновения передними ногами являются индикатором сильного «призыва» к обмену и наблюдаются чаще у акцептора, чем у донора. После насыщения акцептор в свою очередь становится донором. Сходное поведение я наблюдал и у
F. fusca зависит от того, движения какого из партнеров являются более интенсивными. Прикосновения передними ногами являются индикатором сильного «призыва» к обмену и наблюдаются чаще у акцептора, чем у донора. После насыщения акцептор в свою очередь становится донором. Сходное поведение я наблюдал и у ![]() F. rufa. Интересно, что у кызылкумского Cataglyphis foreli oxiana, по нашим данным, наблюдается похожая картина, однако в этом случае движения ног практически не имеют значения, но акцептор во время обмена пищей (особенно вначале) постоянно поглаживает донора нижнечелюстными щупиками по нижней стороне головы.
F. rufa. Интересно, что у кызылкумского Cataglyphis foreli oxiana, по нашим данным, наблюдается похожая картина, однако в этом случае движения ног практически не имеют значения, но акцептор во время обмена пищей (особенно вначале) постоянно поглаживает донора нижнечелюстными щупиками по нижней стороне головы.
Еще один способ обмена информацией был открыт Штегером (Stager, 1931) у рыжих лесных муравьев. Рабочий, нашедший мертвую добычу, начинает возбужденно бегать вокруг нее. Привлеченные этими движениями рабочие, находящиеся поблизости, начинают двигаться сходным образом и устремляются к добыче. Как считал Штегер, приближение рабочих — это результат восприятия движения, контакт же или пахучие вещества не играют роли. Такой способ передачи информации получил название кинопсиса. Струдза (Strudza, 1943a), изучая реакцию рабочих F. pratensis на движущийся диск с полосами, обнаружил, что, когда один муравей начинает двигаться за диском, другие повторяют его движения. Результаты этих опытов исследователь считал экспериментальным доказательством существования кинопсиса.
П. И. Мариковский (1958а, 1965б) описал 14 различных сигналов, преимущественно тактильных, у Camponotus herculeanus. Сходные сигналы, возможно, будут обнаружены и у Formica.
Этот же автор высказывает предположение, что у муравьев имеется биологическая радиосвязь, однако объективных доказательств этому не приводит (Мариковский, 1965б).
Хотя у ряда муравьев, например у наших Myrmica, уже давно известны звуковые органы, роль звуковых сигналов в обмене информацией у муравьев не была подтверждена ни одним серьезным исследованием (Wilson, 1963b).
В последние годы были достигнуты большие успехи в изучении химического обмена информацией; это привело многих исследователей к мнению, что такой способ является у муравьев основным (Wilson, 1963a, b).
Вещества, выделяемые животными и изменяющие поведение других особей того же вида, называются феромонами (эктогормонами, гомотелергонами). Если основная функция феромона пустить в ход, подобно курку, цепь физиологических событий, эффект их действия называется «эффектом толчка» (primer effect) (см. главу V, раздел 4). Другая группа феромонов вызывает «сигнальный эффект» (releaser effect), т.е. сразу же изменяет поведение реципиента и действует, видимо, прямо на центральную нервную систему. Лучше всего изучены относящиеся к последней группе следовые вещества (одмихионы) и торибоны (рис:84).
Для обозначения биологически активных веществ, выделяемых животными и воздействующих на других животных, советским физиологом Я. Д. Киршенблатом был предложен термин «телергоны», а для феромонов — «гомотелергоны». Однако, насколько мне известно, никто из специалистов в этой области последнего термина не применяет. Термин же «феромоны», предложенный Карлсоном и Бутенандтом, широко используется в литературе. Сложную и громоздкую терминологию Киршенблата (в окончательном виде — в Nature, London, v.195, №4844, p.196—197, 1962) нельзя признать удачной. Из выдвинутых им терминов в настоящей работе используется лишь термин «торибоны» (thorybones), так как распространенный в литературе термин «alarm substances» трудно перевести на русский (от английского alarm — переполох, тревога, т.е. дословно — вещества, вызывающие переполох).
Торибоны — это большая и разнообразная группа веществ, видимо в основном терпеноидов, оказывающих различное действие на поведение. У изученных видов они выделяются анальной железой (![]() Dolichoderinae) или чаще мандибулярными железами. В малой дозе это вещество привлекательно для муравьев, а при более высокой концентрации вызывает агрессивное состояние рабочих. Обычно какой-либо муравей, будучи чем-либо возбужден (рис:84), выбрасывает в воздух порцию торибона. При этом виды, у которых эти вещества выделяются мандибулярными железами, совершают резкие движения головой. Привлеченные запахом, другие ра-бочие устремляются к нему, но вблизи концентрация торибона в воздухе повышается, и эти рабочие сами приходят в агрессивное состояние и выбрасывают новые порции торибона. Таким образом, при опасности муравьи быстро собираются вместе. Поскольку торибоны летучи, при прекращении опасности очаг возбуждения затухает. Длительное воздействие большой дозы торибонов стимулирует у Pogonomyrmex badius интенсивные копательные движения, а у Tapinoma sessile вызывает переселение семьи из старого гнезда (все данные по обзорам Wilson, 1963a, b).
Dolichoderinae) или чаще мандибулярными железами. В малой дозе это вещество привлекательно для муравьев, а при более высокой концентрации вызывает агрессивное состояние рабочих. Обычно какой-либо муравей, будучи чем-либо возбужден (рис:84), выбрасывает в воздух порцию торибона. При этом виды, у которых эти вещества выделяются мандибулярными железами, совершают резкие движения головой. Привлеченные запахом, другие ра-бочие устремляются к нему, но вблизи концентрация торибона в воздухе повышается, и эти рабочие сами приходят в агрессивное состояние и выбрасывают новые порции торибона. Таким образом, при опасности муравьи быстро собираются вместе. Поскольку торибоны летучи, при прекращении опасности очаг возбуждения затухает. Длительное воздействие большой дозы торибонов стимулирует у Pogonomyrmex badius интенсивные копательные движения, а у Tapinoma sessile вызывает переселение семьи из старого гнезда (все данные по обзорам Wilson, 1963a, b).
Следовые вещества выделяются различными экскреторными железами (железой Дюфура, ядовитой железой у![]() Myrmicinae, железой Павана у
Myrmicinae, железой Павана у ![]() Dolichoderinae, железами кишечного тракта у
Dolichoderinae, железами кишечного тракта у![]() Formicinae), отверстие которых располагается на конце брюшка. Рабочие, нашедшие пищу, возвращаясь с добычей к гнезду, периодически прикасаются брюшком к субстрату и оставляют пятнышки пахучего следа. Прежде существовало мнение, что у всех видов по возвращении в гнездо «фуражиры» каким-либо образом сообщают другим муравьям о нахождении пищи и побуждают их выходить из гнезда. Однако исследования Вильсона с Solenopsis saevissima показали, что для этого достаточно одних следовых веществ. Муравей, обнаруживший след (у этого вида следовые вещества быстро испаряются), сразу же направляется по нему. Количество информации, передаваемой таким образом, приблизительно равно количеству информации пчелиного «танца» (Wilson, 1963a).
Formicinae), отверстие которых располагается на конце брюшка. Рабочие, нашедшие пищу, возвращаясь с добычей к гнезду, периодически прикасаются брюшком к субстрату и оставляют пятнышки пахучего следа. Прежде существовало мнение, что у всех видов по возвращении в гнездо «фуражиры» каким-либо образом сообщают другим муравьям о нахождении пищи и побуждают их выходить из гнезда. Однако исследования Вильсона с Solenopsis saevissima показали, что для этого достаточно одних следовых веществ. Муравей, обнаруживший след (у этого вида следовые вещества быстро испаряются), сразу же направляется по нему. Количество информации, передаваемой таким образом, приблизительно равно количеству информации пчелиного «танца» (Wilson, 1963a).
Следовые вещества у Formica неизвестны. Имеющиеся в литературе указания (Щербаков, 1953; Goetsch, 1953), что рыжие лесные муравьи метят тропу муравьиной кислотой, ни на чем не основаны и не проверены экспериментально. Штумпер (Stumper, 1959а) обнаружил новый компонент в составе муравьиного яда и предположил, что это следовое вещество, но опять же никаких доказательств этому не приводится. Кстати, у всех изученных видов подсемейства![]() Formicinae —
Formicinae — ![]() Lasius fuliginosus, Paratrechina longicornis, Myrmelachista ramulorum (Blum, Wilson, 1964) — следовые вещества выделяются из задней кишки.
Lasius fuliginosus, Paratrechina longicornis, Myrmelachista ramulorum (Blum, Wilson, 1964) — следовые вещества выделяются из задней кишки.
Полученные нами данные позволяют сделать вывод, что у Formica следовые вещества либо вообще отсутствуют, либо играют очень незначительную роль. То же, очевидно, относится к Cataglyphis, Camponotus и многим другим родам. Предположение Вильсона, что информация о наличии пищи вне гнезда передается только следовыми феромонами, является, видимо, преувеличением.
Остановимся подробнее на некоторых наблюдениях над Formica, проделанных осенью 1965 г. Если нанести пипеткой на лист бумаги след вытяжкой из брюшка муравьев, имеющих следовые феромоны, то выпущенные на лист муравьи того же или близкого вида, обнаружив искусственную тропу, начинают двигаться по ней (Wilson, 1963а). Опыты с водной и эфирной вытяжками ![]() F. rufa и F. pratensis не дали положительных результатов. Точно так же, если положить на тропу муравьев, следовые феромоны которых хорошо известны, например, Tapinoma, лист бумаги, то через некоторое время (в наших опытах с T. karavaievi — менее суток) рабочие делают четкую пахучую тропу, по которой следуют выпущенные на лист особи того же вида. Ничего подобного ни у
F. rufa и F. pratensis не дали положительных результатов. Точно так же, если положить на тропу муравьев, следовые феромоны которых хорошо известны, например, Tapinoma, лист бумаги, то через некоторое время (в наших опытах с T. karavaievi — менее суток) рабочие делают четкую пахучую тропу, по которой следуют выпущенные на лист особи того же вида. Ничего подобного ни у ![]() F. rufa, ни у F. pratensis нам не удалось обнаружить.
F. rufa, ни у F. pratensis нам не удалось обнаружить.
На тропах лугового муравья мы вкапывали вровень с землей круги из фанеры и жести. В аналогичных условиях T. karavaievi, Pheidole pallidula arenarum и другие муравьи, имеющие следовые феромоны, спустя, по крайней мере, сутки начинают ходить поперек круга по кратчайшему расстоянию. В то же время, рабочие лугового муравья, четкие и узкие тропы которого хорошо известны, даже спустя полторы недели предпочитали ходить по краю круга, где имелись хорошие наземные ориентиры. В естественных условиях нам неоднократно приходилось наблюдать, как тропы этого муравья, выходя на открытые участки (дорога, песчаная площадка и т.д.), утрачивают свою четкость и правильность: муравьи начинают сильно уклоняться от прямого направления, несмотря на то, что тропа через этот участок существует уже несколько лет.
Как доказательство отсутствия следовых веществ у Formica можно привести и один из опытов со светокомпасной ориентацией ![]() F. sanguinea. После того, как «фуражир» в результате дрессировки перестал реагировать на переключение света (см. предыдущий раздел главы), на 1 час 20 мин. была включена основная лампа. Все это время муравей носил куколок в гнездо. Затем, когда он находился на кормушке, вновь было произведено переключение света под углом 90°. Отклонения от прямого пути появились вновь и отмечались в течение первых четырех рейсов (рис:82/8/). Продолжительность рейсов увеличилась в 2,5—7 раз. Затем муравей вновь перестал реагировать на переключение света (рис:83). Если бы путь муравья отмечался следовыми феромонами, результат должен был бы быть обратным, т.е. чем больше времени прошло с начала опыта, тем четче должен быть пахучий след и, следовательно, слабее реакция на переключение света.
F. sanguinea. После того, как «фуражир» в результате дрессировки перестал реагировать на переключение света (см. предыдущий раздел главы), на 1 час 20 мин. была включена основная лампа. Все это время муравей носил куколок в гнездо. Затем, когда он находился на кормушке, вновь было произведено переключение света под углом 90°. Отклонения от прямого пути появились вновь и отмечались в течение первых четырех рейсов (рис:82/8/). Продолжительность рейсов увеличилась в 2,5—7 раз. Затем муравей вновь перестал реагировать на переключение света (рис:83). Если бы путь муравья отмечался следовыми феромонами, результат должен был бы быть обратным, т.е. чем больше времени прошло с начала опыта, тем четче должен быть пахучий след и, следовательно, слабее реакция на переключение света.
Таким образом, вопрос о том, как виды рода Formica передают информацию о наличии пищи на кормовом участке, остается пока неясным.
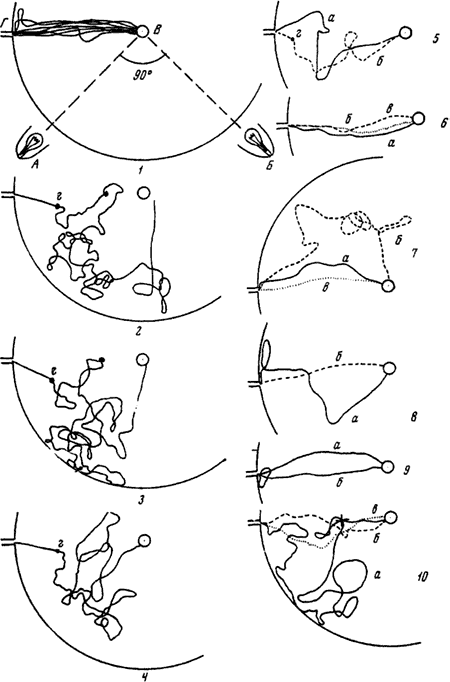
Рис. 82. Реакция рабочих
На рисунках 1—9 изображен путь рабочего от кормушки с куколками (В) до входа в гнездо (В) в разных опытах, когда были включены либо лампа А, либо лампа Б. На рисунках 1, 7, 8,9б показан путь одного и того же рабочего, когда была включена лампа А, а на рисунках 2—6, 9а — путь того же муравья, когда он двигался от кормушки к входу при свете лампы Б. г (на всех рисунках) — точки, из которых рабочие замечали отверстие входа в гнездо. На рисунке 10 изображен путь другого рабочего. В этом опыте переключение света производилось в тот момент, когда муравей находился в точке, отмеченной крестом: а — 2-й рейс после переключения (самый продолжительный); б — 10-й рейс после переключения; в — первый рейс после того, как перестали переключать свет
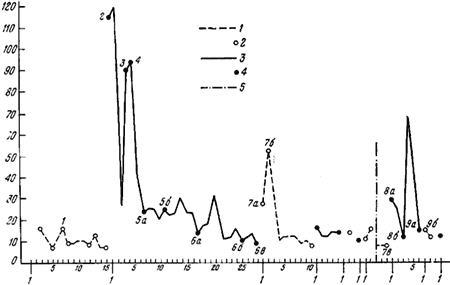
Рис. 83. Реакция рабочих
На горизонтальной оси — порядок опытов, на вертикальной — продолжительность рейсов в секундах. На этом рисунке изображены результаты того же опыта, что и на рис.82/1—9/; цифры на графиках соответствуют номерам этого рисунка; 1,2 — включена только лампа А; 3, 4 — дорогу от кормушки к гнезду муравей находит при свете лампы Б; 5 — перерыв на 1 час. 20 мин. (все время была включена лампа А)
 Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant
Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелец
Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелециндивидуальное или групповое привлечение пассивных фуражиров активными фуражирами для конкретных полей в конкретном месте.
выполнение муравьем в течение длительного времени определенных функций в семье. Если функция связана только с возрастом - возрастной полиэтизм; если функция выполняется постоянно - постоянный, или «кастовый», полиэтизм.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
наука изучающая муравьев, часть энтомологии. Мирмекологи - ученые изучающие муравьев.
(от трофо… и греч. allaxis — обмен) передача пищи и гормональных веществ от особи к особи путем кормления содержимым зобика, желудка или слизывания выпота с поверхности тела. Играет огромную роль в передаче информации между муравьями.
 Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant
Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant(лат. acceptor - приемник) - принимающий на себя.
 Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравейксеробионт
восприятие муравьями в качестве сигнала характерных движений других особей одной общины («язык поз»).
Всего 965 видов.
(греч. φέρω — «нести» + ορμόνη — «гормон») — летучие вещества, управляющие поведенческими реакциями, процессами развития, а также многими процессами, связанными с социальным поведением и размножением. Феромоны модифицируют поведение, физиологическое и эмоциональное состояние или метаболизм других особей того же вида.
Небольшое наиболее архаичное подсемейство муравьев. Распространено по всему земному шару, преимущественно в тропиках. Насчитывает более 230 видов.
(лат. mandibule — челюсть, от mando — жую, грызу), первая пара челюстей у ракообразных, многоножек и насекомых. Синоним: жвала.
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
Myrmicinae — самое большое подсемейство муравьев, включающее более 2 тыс. видов, относящихся более чем к 150 родам.
Одно из крупных подсемейств, насчитывает около 1,5 тыс. видов, относящихся к 44 родам. Широко распространены на всем земном шаре, довольно обычны в тропиках и занимают доминирующее положение в умеренных широтах.
надсемейство летающих насекомых отряда перепончатокрылых, родственное осам и муравьям. Существует около 20 тысяч видов пчёл. Распространены на всех континентах, кроме Антарктиды. Пчёлы питаются нектаром и пыльцой, используя нектар главным образом в качестве источника энергии, а пыльцу для получения белков и других питательных веществ.
 Lasius fuliginosus (Latreille, 1798) — пахучий муравей-древоточец, jet ants
Lasius fuliginosus (Latreille, 1798) — пахучий муравей-древоточец, jet antsВсего 65 видов.
Всего 62 вида.
гигрофил, галобионт
Старое название: Pheidole arenarum Ruzsky
участок, используемый муравьями для добычи пищи.






