






Все виды рода Formica имеют смешанное питание. Рацион взрослых особей составляют в основном углеводы, а личинок — белки. Но даже для поддержания постоянного количества особей в гнезде при отсутствии хищников, поедающих рабочих муравьев в природе, все же необходимо постоянное количество белковой пищи. Канадский мирмеколог Эйр (Ayre, 1960) провел следующий опыт: в каждый из 8 формикариев было помещено по 3000 рабочих и «царица» F. polyctena, взятых из одного гнезда. Все экспериментальные семьи содержались при температуре 26—28° и получали одинаковое (избыточное) количество углеводной пищи (100 г меда на 100 мл воды), но различное количество белковой (0,50, 100, 200, 400, 800, 1600 мг личинок мух в сутки). Через 5 месяцев было подсчитано количество особей в каждой из экспериментальных семей. Результаты этого интересного опыта изображены на рис:79. В гнезде, не получавшем белковой пищи, население уменьшилось почти вдвое, а в гнезде, получавшем 1600 мг в сутки, — увеличилось вдвое. Количество особей остается постоянным, если семья будет получать около 220 мг личинок, т. е. 7,3-10-5 г на особь в сутки. Сходные данные были получены, независимо от Эйра, Ланге (Lange, 1960с). Естественно, что в полевые условия эти данные можно переносить лишь как минимальные, так как в опытах не учитывается количество пищи, необходимое для выведения половых особей и пополнения естественного отпада.

Рис. 79. Изменения в количестве населения семей F.polyctena из 3000 рабочих и самки при кормлении в течение 5 месяцев различным количеством белковой пищи (Ayre, 1960)
Количественное соотношение раз-личных пищевых компонентов у рыжих лесных муравьев зависит от местных условий, состояния гнезда и вида муравьев и меняется в течение сезона. Так, например, Г. А. Милованова (1963) наблюдала у F. polyctena в Приокско-террасном заповеднике следующую смену: весной эти муравьи питаются в основном березовым соком, в конце весны и в первой половине лета основу их пищи составляют насекомые, а со второй половины лета и до ухода на зимовку эти муравьи питаются в основном падью тлей. Сходную закономерность на-блюдал Эйр (Ayre, 1957) у F. subnitens Creight. в Канаде. Вообще переход от преимущественно белкового питания к углеводному во второй половине лета, по-видимому, вообще характерен для этих муравьев.
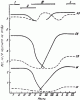
Рис. 80. Сезонные изменения абсолютного и сухого веса и содержания жира у рабочих F. polyctena (Kirchner, 1934)
АВ — абсолютный вес; СВ — сухой вес; Ж — содержание жира; I — зимовка; II — выведение половых особей; III — выведение рабочих; сплошная линия — молодые рабочие; пунктирная линия — старые рабочие
Эти наблюдения согласуются с экспериментальными данными. Так, Ланге (Lange, 1962) показал, что даже в экспериментальных условиях, при наличии постоянного количества пищи (личинки Musca domestica), у ![]() F. rufa и F. polyctena существует ритмика охотничей активности в течение длительных периодов времени. Эти фазы активности связаны с фазами выведения молоди. Во время появления в гнездах большого количества личинок старших возрастов количество уносимых в гнездо личинок мух увеличивается, а после их исчезновения — уменьшается.
F. rufa и F. polyctena существует ритмика охотничей активности в течение длительных периодов времени. Эти фазы активности связаны с фазами выведения молоди. Во время появления в гнездах большого количества личинок старших возрастов количество уносимых в гнездо личинок мух увеличивается, а после их исчезновения — уменьшается.
Данные, полученные Кирхнером (Kirchner, 1961, 1964), свидетельствуют о том, что абсолютный вес рабочих муравьев ![]() F. rufa, F. polyctena, F. pratensis и F. exsecta значительно уменьшается в течение 4 недель после зимнего покоя, в начале лета увеличивается, уменьшается в конце лета и резко возрастает осенью, после окончания выведения молоди (рис:80). Сухой вес уменьшается весной, остается постоянным в течение лета и снова повышается осенью. Количество (абсолютное и относительное) жира сильно уменьшается весной, достигает минимума летом и снова возрастает осенью. У молодых рабочих как абсолютный вес, так и содержание жира выше. Это указывает на то, что именно они воспитывают молодь в гнезде (глава V).
F. rufa, F. polyctena, F. pratensis и F. exsecta значительно уменьшается в течение 4 недель после зимнего покоя, в начале лета увеличивается, уменьшается в конце лета и резко возрастает осенью, после окончания выведения молоди (рис:80). Сухой вес уменьшается весной, остается постоянным в течение лета и снова повышается осенью. Количество (абсолютное и относительное) жира сильно уменьшается весной, достигает минимума летом и снова возрастает осенью. У молодых рабочих как абсолютный вес, так и содержание жира выше. Это указывает на то, что именно они воспитывают молодь в гнезде (глава V).
Исходя из изложенных выше фактов, а также данных о динамике выведения молоди в гнезде (глава V), можно построить следующую схему: весной, после выхода из зимовки, рыжие лесные муравьи начинают воспитывать половых особей. При этом они расходуют в основном запасы, накопленные ими осенью. Другие насекомые в это время малоактивны, а тли еще не образуют колоний. Потери в весе лишь немного восполняются за счет питания соком деревьев. Сразу же после вылета крылатых в гнездах начинается массовое выведение рабочих, которое продолжается первую половину лета. Поэтому в этот период муравьи главным образом добывают пищу для личинок, т. е. охотятся на различных мелких беспозвоночных. Колонии тлей в начале лета еще невелики, и поэтому значительную роль в питании взрослых муравьев играют эласмосомы семян (Oekland, 1932).
К началу осени выведение молоди в гнездах в основном заканчивается и одновременно сильно разрастаются колонии тлей. В это время муравьи питаются в основном углеводами, которые перерабатываются ими в жировое тело, используемое затем при выведении весеннего поколения крылатых.
Накоплено уже много данных о составе пищи Formica и соотношении ее различных компонентов. Однако в результате неточностей при определении муравьев большинство этих данных не может быть использовано.
Приведем некоторые литературные данные о количестве пищи, потребляемой одной семьей рыжего лесного муравья, не уточняя, однако, видового определения.
Велленштайн (Wellenstein, 1959, 1955) на основе 25-летних наблюдений дает следующие средние цифры: активность рыжих лесных муравьев начинается в Средней Европе в конце марта и кончается в октябре. За это время одна средняя семья собирает 3—8 млн. насекомых, 200 л углеводов (преимущественно в виде пади тлей) и 40—60 тыс. семян. Среднее весовое соотношение различных компонентов: 62% — падь, 33% — насекомые и другие беспозвоночные, 4,5% — сок растений, 0,3% грибы и падаль и 0,2% — семена.
Холт (Holt, 1955) вычислил, что в июле средняя продолжительность фуражировочной экспедиции рабочего составляет около 2 час. Исходя из этого времени и количества рабочих, покинувших гнездо, автор установил, что вне гнезда работает около 70 тыс. фуражиров, весящих около 700 г. За 14 час. активности они приносят 820 г пищи, 44% которой составляет падь. Каждый рабочий, таким образом, приносит за сутки добычу, превышающую его собственный вес.
По данным большинства авторов, рабочие одного гнезда рыжего лесного муравья приносят в гнездо в течение суток 20—100 тыс. убитых насекомых (Forel, 1874; Eidmann, 1930; Б. А. Смирнов, 1962а, б, 1963), но это, видимо, максимальные цифры.
Результаты исследований Экленда (Oekland, 1932) не согласуются с изложенным выше. По-видимому, это объясняется тем, что он имел дело не с рыжими лесными муравьями, а с луговыми муравьями (F. pratensis). 100 тыс. рабочих, приходящих в гнездо в течение 20 час, собирают 50—60 г пади и только 2,5 г (сухой вес) белковой пищи. В течение первой половины лета эти муравьи приносят около 3000 семян.
Относительное и абсолютное количество пади в питании разных видов муравьев различно. У F. pratensis она является основным компонентом, а насекомых они уничтожают мало (Gosswald, 1941c и др.). ![]() F. rufa (средняя и большая расы рыжего лесного муравья) добывают сравнительно больше насекомых. Наиболее активным хищником является F. polyctena (Gosswald, 1941b, c, 1944,1954b и др.). Одна семья
F. rufa (средняя и большая расы рыжего лесного муравья) добывают сравнительно больше насекомых. Наиболее активным хищником является F. polyctena (Gosswald, 1941b, c, 1944,1954b и др.). Одна семья ![]() F. rufa собирает в течение года 450—500 кг пади, или 90—100 кг сахара, а F. polyctena (малая раса) — 290—320 кг или 60—65 кг сахара (Zoebelein, 1956).
F. rufa собирает в течение года 450—500 кг пади, или 90—100 кг сахара, а F. polyctena (малая раса) — 290—320 кг или 60—65 кг сахара (Zoebelein, 1956).
Количество пищи, которую муравьи добывают в течение суток, в большой степени зависит от их активности. На суточную активность «фуражиров» влияют многие факторы (температура воздуха и почвы, влажность, осадки, освещенность и т. д.), но определяющим фактором в наших условиях является температура (Wellenstein, 1954а, 1957а; Schwenke, 1957; R. Huber, 1961). Температура влияет на активность муравьев двояко. С одной стороны, скорость движения рабочих находится в прямой зависимости от температуры воздуха (Strudza, 1935; Stumper, 1922а; Ayre, 1958b; Holt, 1955), и поэтому при понижении температуры продолжительность экспедиции каждого «фуражира» увеличивается, а следовательно, производительность падает (Holt, l. c.). С другой стороны, в зависимости от температуры из гнезда выходит на фуражировку различное количество рабочих.
Вопрос о том, существует ли у муравьев эндогенный ритм активности, до сих пор остается открытым. Имеются косвенные доказательства о наличии такого ритма у муравьев-листорезов (Atta) (Hodgson, 1955). В то же время у Iridomyrmex humilis и Crematogaster scutellaris (Мак Клускей, Солье, по Wilson, 1963с) фуражировка экзогенно определяется температурой. В отношении Formica большинство авторов считает, что ритм активности этих муравьев определяется климатическими факторами (Bruns, 1945а; Dobrzanski, 1956; Reichle, 1943).
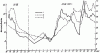
Рис. 81. График суточной активности F. pratensis (Воронежский заповедник, 29 — 30.VII 1955 г.)
1 — интенсивность ухода; 2 — интенсивность прихода; 3 — температура на поверхности купола
29 и 30 мая 1965 г. в Воронежском заповеднике автор провел суточный учет активности рабочих гнезда F. pratensis, расположенного на поляне 40-летнего сосняка-зеленомошника.
Методика наблюдений была следующей. На расстоянии 1,5 м от муравейника на одной из дорожек наносилась контрольная линия и отмечалось количество муравьев, проходящих через нее в течение 3 мин. к муравейнику и от него. Количество муравьев, прошедших через контрольную линию за одну минуту от муравейника, мы обозначили как интенсивность ухода, к муравейнику — как интенсивность прихода, количество муравьев с добычей — как интенсивность охоты. Одновременно отмечалась температура на поверхности муравейника и температура воздуха в тени. Измерения проводились через каждые 15 мин., а затем была сделана обработка данных методом скользящей кривой на 5 точек.
График суточной активности приводится на рис:81. Из этого графика можно сделать следующие выводы:
1. В первую половину дня интенсивность ухода выше интенсивности прихода, во вторую половину дня — соотношения обратные, причем перед заходом солнца интенсивность прихода особенно повышается.
2. Оптимальная температура лежит около +20°, и дальнейшее повышение температуры действует угнетающе, в результате чего в середине дня наблюдается спад активности.
3. Интенсивность ухода гораздо сильнее зависит от колебания температуры поверхности гнезда, чем интенсивность прихода.
4. Хотя интенсивность прихода гораздо выше в вечерние часы, интенсивность охоты выше утром и днем (10—12 час.).
Сопоставление интенсивности прихода и охоты показывает, что в течение утреннего пика активности добычу несет постоянное число муравьев (около 24%), а во время вечернего пика наблюдается большой разброс. По-видимому, это связано с тем, что во второй половине дня в гнездо возвращается много «пустых» рабочих и количество рабочих с пищей определяется случайными причинами.
Относительно ![]() F. rufa и F. polyctena большинство авторов также приходит к выводу, что оптимальная температура фуражировки лежит около 20°, и в жаркие дни в середине дня наблюдается спад активности (Wellenstein, 1953; Otto, 1962). В теплые ночи муравьи также могут быть активны (Wellenstein, 1957а).
F. rufa и F. polyctena большинство авторов также приходит к выводу, что оптимальная температура фуражировки лежит около 20°, и в жаркие дни в середине дня наблюдается спад активности (Wellenstein, 1953; Otto, 1962). В теплые ночи муравьи также могут быть активны (Wellenstein, 1957а).
Оптимальные температуры могут варьировать даже для одного гнезда в зависимости от условий, но во всех случаях фуражировку муравьи начинают при температуре не ниже 7—10°.
Так как муравьи нападают преимущественно на движущихся насекомых, наиболее интенсивно они охотятся на тех насекомых, температурный оптимум которых лежит также около 20°, и активных в первую половину дня.
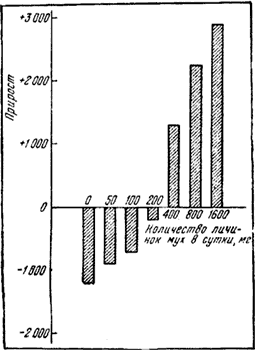
Рис. 79. Изменения в количестве населения семей F.polyctena из 3000 рабочих и самки при кормлении в течение 5 месяцев различным количеством белковой пищи (Ayre, 1960)
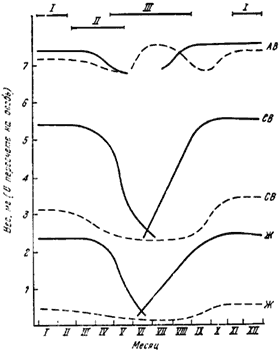
Рис. 80. Сезонные изменения абсолютного и сухого веса и содержания жира у рабочих F. polyctena (Kirchner, 1934)
АВ — абсолютный вес; СВ — сухой вес; Ж — содержание жира; I — зимовка; II — выведение половых особей; III — выведение рабочих; сплошная линия — молодые рабочие; пунктирная линия — старые рабочие
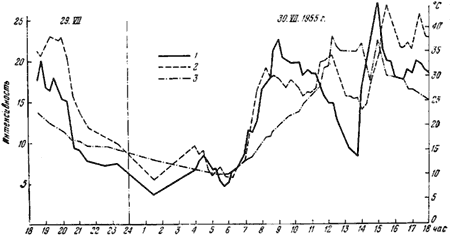
Рис. 81. График суточной активности F. pratensis (Воронежский заповедник, 29 — 30.VII 1955 г.)
1 — интенсивность ухода; 2 — интенсивность прихода; 3 — температура на поверхности купола
наука изучающая муравьев, часть энтомологии. Мирмекологи - ученые изучающие муравьев.
искусственный муравейник.
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
сладкая густая жидкость, выделяемая тлями, червецами и другими насекомыми, питающимися соком растений. Медоносная пчела собирает падь с листьев и перерабатывает в падевый мед.
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
 Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравейфункциональное образование, состоящее из материнской и дочерних семей, поддерживающих регулярные родственные отношения
животные, не имеющие позвоночника: простейшие, губки, кишечнополостные, черви, моллюски, членистоногие, иглокожие и др.; примерно 3/4 видов всех животных.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
Старое название: Iridomyrmex humilis






