






При поисках добычи и при возвращении в гнездо муравьи должны хорошо ориентироваться на местности. Основным способом ориентации у Formica является светокомпасная ориентация.
Начало исследованиям в этом направлении было положено классическими опытами известного английского естествоиспытателя Леббока (1898). Этот исследователь помещал на арену искусственного гнезда садового муравья (![]() Lasius niger) чашечку с куколками. С одной стороны гнезда ставилась горящая свеча. После того, как муравьи запоминали дорогу и начинали ходить с куколками почти по прямой, свеча переставлялась на другую сторону арены. После этого муравьи меняли направление движения на противоположное и с большим трудом находили вход в гнездо.
Lasius niger) чашечку с куколками. С одной стороны гнезда ставилась горящая свеча. После того, как муравьи запоминали дорогу и начинали ходить с куколками почти по прямой, свеча переставлялась на другую сторону арены. После этого муравьи меняли направление движения на противоположное и с большим трудом находили вход в гнездо.
Второй классический опыт (Brun, 1914), вошедший во все учебники энтомологии, также был поставлен впервые на садовом муравье. Если муравья, который движется к гнезду в естественных условиях, прикрыть темным колпаком и спустя 1—2 часа выпустить, то он начинает двигаться также по прямой, но под некоторым углом, равным углу, на который за время опыта переместилось солнце к первоначальному направлению. Впоследствии этот опыт неоднократно повторяли на других видах му-равьев.
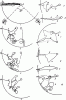
Рис. 82. Реакция рабочих
На рисунках 1—9 изображен путь рабочего от кормушки с куколками (В) до входа в гнездо (В) в разных опытах, когда были включены либо лампа А, либо лампа Б. На рисунках 1, 7, 8,9б показан путь одного и того же рабочего, когда была включена лампа А, а на рисунках 2—6, 9а — путь того же муравья, когда он двигался от кормушки к входу при свете лампы Б. г (на всех рисунках) — точки, из которых рабочие замечали отверстие входа в гнездо. На рисунке 10 изображен путь другого рабочего. В этом опыте переключение света производилось в тот момент, когда муравей находился в точке, отмеченной крестом: а — 2-й рейс после переключения (самый продолжительный); б — 10-й рейс после переключения; в — первый рейс после того, как перестали переключать свет
В начале нынешнего века против светокомпасной ориентации у муравьев выдвигались серьезные возражения. Дело в том, что муравьи продолжают двигаться к гнезду по прямой и в том случае, когда они не видят солнца. Это заставило Корнеца (обзор работ — Cornetz, 1914) выдвинуть гипотезу о наличии у них «внутреннего компасного чувства». Санчи (обзор работ — Santschi, 1923) отрицал наличие такого чувства у муравьев и для объяснения этих случаев высказал предположение, что муравьи способны видеть степень освещенности поверхности почвы, но эти возражения трудно было принимать всерьез, поскольку, например, в лесу, как справедливо отмечал Корнец (l. c.), свет, проходя через листву, создает на почве сложную мозаику световых пятен различной освещенности, что, однако, не сказывается на способности муравьев ориентироваться в этих условиях. Объяснение было получено недавно, когда Воулес (Vowles, 1950) на примере ![]() Myrmica rubra и M. ruginodis показал, что муравьи, подобно пчелам, способны видеть направление поляризации света.
Myrmica rubra и M. ruginodis показал, что муравьи, подобно пчелам, способны видеть направление поляризации света.
Наиболее полные исследования, посвященные ориентации муравьев, были проведены около 10 лет назад Яндером (Jander, 1957) в ФРГ. В качестве объекта были выбраны рыжие лесные муравьи. Было убедительно показано, что у этих муравьев имеется три основных способа ориентации 1) по отчетливому источнику света (солнце или луна в естественных условиях), 2) по направлению колебаний света (поляризация голубого неба в естественных условиях) и 3) по наземным ориентирам. Первый способ используется тогда, когда муравьи видят солнце или луну, второй — в тени или если солнце скрыто за облаками, но видны участки голубого неба, а третий — в пасмурную погоду. В искусственных условиях поворот муравьев, движущихся в определенном направлении, может быть вызван перемещением (переключением) источника света, если свет падает на арену гнезда несколько сбоку, поворотом листа поляроидной пленки, которым прикрыт источник света, расположенный над ареной, или перемещением макетов деревьев, если арена освещена рассеянным светом.
В естественных условиях молодые рабочие совершают первые фуражировочные экскурсии поблизости от гнезда и лишь после ознакомления с внешними ориентирами начинают удаляться от него (Brun, 1914).
Наши исследования показали, что в условиях опыта муравьи быстро приспосабливаются к правильной светокомпасной ориентации даже в условиях, с которыми они никогда не встречаются в природе.
На арену искусственного гнезда ![]() F. sanguinea выставлялась чашка с куколками F. pratensis или F. exsecta. Единственным ориентиром для муравьев была лампа. Спустя некоторое время, когда муравьи начали таскать куколок до гнезда почти по прямой, основная лампа (А) выключалась и включалась дополнительная, свет от которой падал на арену под углом 90° к первоначальному направлению (рис:82/1/, лампы А и Б). Муравьи, взявшие куколок, начинали идти не к гнезду, а под углом 90° к первоначальному направлению. Продолжительность рейса возрастала в 10 раз. Постепенно путь все более приближался к прямой и после 12 рейсов восстанавливалось нормальное движение. Этот опыт является повторением уже известных опытов Леббока, Яндера и др. (l. c.). Следует только отметить, что в опытах Яндера рыжие лесные муравьи приспосабливались к изменяющимся условиям освещения иногда лишь на пятые сутки. Аналогичным образом в наших опытах рабочие
F. sanguinea выставлялась чашка с куколками F. pratensis или F. exsecta. Единственным ориентиром для муравьев была лампа. Спустя некоторое время, когда муравьи начали таскать куколок до гнезда почти по прямой, основная лампа (А) выключалась и включалась дополнительная, свет от которой падал на арену под углом 90° к первоначальному направлению (рис:82/1/, лампы А и Б). Муравьи, взявшие куколок, начинали идти не к гнезду, а под углом 90° к первоначальному направлению. Продолжительность рейса возрастала в 10 раз. Постепенно путь все более приближался к прямой и после 12 рейсов восстанавливалось нормальное движение. Этот опыт является повторением уже известных опытов Леббока, Яндера и др. (l. c.). Следует только отметить, что в опытах Яндера рыжие лесные муравьи приспосабливались к изменяющимся условиям освещения иногда лишь на пятые сутки. Аналогичным образом в наших опытах рабочие ![]() F. rufa в течение первых суток двигались от кормушки к входу в гнездо в пределах сектора 90°. На вторые сутки этот угол уменьшился до 45°. Дальше опыт не проводился. Все эти данные указывают на значительно более высокую способность к ориентации у
F. rufa в течение первых суток двигались от кормушки к входу в гнездо в пределах сектора 90°. На вторые сутки этот угол уменьшился до 45°. Дальше опыт не проводился. Все эти данные указывают на значительно более высокую способность к ориентации у ![]() F. sanguinea, нежели у
F. sanguinea, нежели у ![]() F. rufa.
F. rufa.
Вернемся к опытам с ![]() F. sanguinea. В дальнейшем опыт был усложнен таким образом, чтобы поставить муравьев в условия, с которыми они никогда не сталкивались в природе. Когда муравей двигался к кормушке, оставалась включенной лампа А, по которой он ориентировался прежде. В тот момент, когда он находился на кормушке и брал куколку, свет переключался, и загоралась лампа Б. После того, как муравей скрывался с куколкой во входе в гнездо, снова включалась лампа А. Было поставлено несколько серий таких опытов, но каждый раз лишь с одним меченым муравьем, и результаты всегда отличались лишь незначительно. На рис:82 показано, как от рейса к рейсу менялся путь муравья, а на рис:83 — как менялась продолжительность рейсов в том же опыте.
F. sanguinea. В дальнейшем опыт был усложнен таким образом, чтобы поставить муравьев в условия, с которыми они никогда не сталкивались в природе. Когда муравей двигался к кормушке, оставалась включенной лампа А, по которой он ориентировался прежде. В тот момент, когда он находился на кормушке и брал куколку, свет переключался, и загоралась лампа Б. После того, как муравей скрывался с куколкой во входе в гнездо, снова включалась лампа А. Было поставлено несколько серий таких опытов, но каждый раз лишь с одним меченым муравьем, и результаты всегда отличались лишь незначительно. На рис:82 показано, как от рейса к рейсу менялся путь муравья, а на рис:83 — как менялась продолжительность рейсов в том же опыте.
После переключения света муравьи начинали двигаться от кормушки, как и в предыдущих опытах, под углом 90° к первоначальному направлению (рис:82/2,3/). Продолжительность рейсов сразу же увеличивалась в 10—60 раз. К 5—6-му рейсу муравьи начинали после переключения света отклоняться лишь на 45° (в начале пути), а время рейсов сокращалось (рис:82/4/; рис:83). После 21—25 рейсов, т. е. приблизительно вдвое медленнее, чем в предыдущем опыте, восстанавливалось нормальное движение (рис:82/5,6/).
После того, как муравьи начинали правильно ориентироваться, несмотря на переключение ламп, мы переставали выключать лампу А. Муравьи продолжали «делать поправку» на переключение и в результате начинали отклоняться в противоположную сторону (рис:82/7/). Однако дорогу к гнезду они находили быстрее, чем при первом изменении обстановки, а продолжительность рейсов увеличивалась всего в 3—5 раз. Через 5—8 рейсов муравьи снова начинали ходить почти по прямой. После этого муравьи вообще переставали реагировать на переключение света и продолжали ходить почти по прямой, независимо от того, какая из ламп была включена. Если свет переключать не в тот момент, когда муравьи находятся на кормушке, а тогда, когда они уже движутся по направлению к гнезду, приспосабливание происходит во много раз быстрее: после 6—7 рейсов снова восстанавливается нормальное движение, и муравьи вообще перестают реагировать на переключение (рис:82/10/).
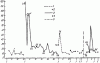
Рис. 83. Реакция рабочих
На горизонтальной оси — порядок опытов, на вертикальной — продолжительность рейсов в секундах. На этом рисунке изображены результаты того же опыта, что и на рис.82/1—9/; цифры на графиках соответствуют номерам этого рисунка; 1,2 — включена только лампа А; 3, 4 — дорогу от кормушки к гнезду муравей находит при свете лампы Б; 5 — перерыв на 1 час. 20 мин. (все время была включена лампа А)
Единственное приемлемое объяснение /О роли пахучих следовых веществ у этих муравьев см. следующий раздел главы/ результатов этого опыта, на наш взгляд, заключается в том, что муравьи после дрессировки начинают воспринимать переключение света, с которым они никогда не встречались и не могли встретиться в природе, и делать соответствующие поправки при ориентации.
Для того, чтобы правильно ориентироваться по солнцу, необходимо постоянно вносить поправки на его перемещение. У муравьев, как и у многих других животных, имеется так называемое «чувство времени». Наличие его доказывается двумя путями. С одной стороны, как показали наблюдения Яндера (Jander, 1957), рыжие лесные муравьи, задержавшиеся на арене искусственного гнезда, освещенного неподвижным источником света, при возвращении в гнездо «делают поправку» на перемещение солнца, т.е. уклоняются от правильного пути под углом, равным тому углу, на который за это время должно было бы переместиться солнце. С другой стороны, известно, что у муравьев можно выработать условный рефлекс на время и приучить их приходить к кормушкам в определенные часы (Шовен, 1953). Рефлекс может сохраняться до 5 суток. Если усыпить муравьев на несколько часов хлороформом или эфиром, условные рефлексы сохраняются (Kalmus, 1934, по Шовену, 1953). Однако введение с пищей в организм муравьев различных химических веществ, влияющих на скорость течения процессов жизнедеятельности, изменяет ритмику. Так, введение иодированного тиреоглобулина, вызывающего повышение температуры тела, заставляет муравьев появляться у кормушек раньше положенного срока, а введение эухинина — запаздывать. Силициловая кислота и красный фосфор вызывают ускорение, а мышьяковая кислота в малых дозах — запаздывание, а в больших — ускорение физиологических процессов (Grabensberger, 1934a, b). Таким образом, «чувство времени» обусловливается ходом физиологических процессов в организме муравьев. Ряд авторов (Reichle, 1943; Dobrzanski, 1956), однако, отрицают наличие чувства времени у муравьев.
Пьеро (Pieron, 1904) считал, что у муравьев имеется также «мускульное чувство времени». При этом он основывался на следующем опыте: муравей, идущий к гнезду, осторожно пересаживался на другую дорожку. После этого он продолжал идти в том же направлении. Но, пройдя то расстояние, которое ему оставалось пройти до гнезда, муравей начинал беспорядочно бегать. Однако при разных усилиях (например, возвращение с грузом или без него, случайные препятствия на пути) муравьи должны были бы ошибаться, если бы они действительно «руководствовались» количеством затраченной мышечной энергии (Cornetz, 1914).
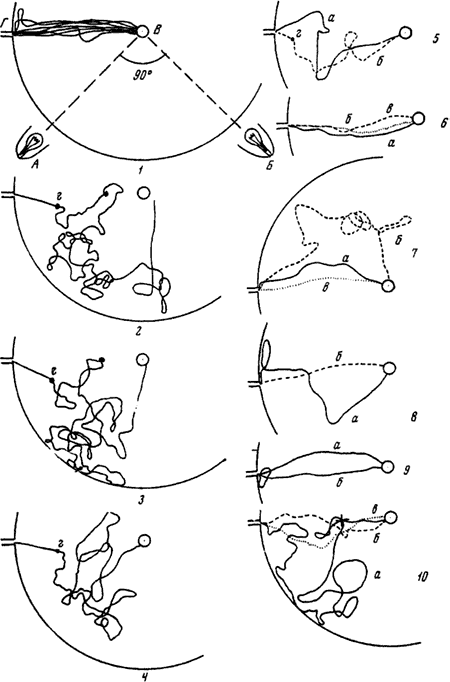
Рис. 82. Реакция рабочих
На рисунках 1—9 изображен путь рабочего от кормушки с куколками (В) до входа в гнездо (В) в разных опытах, когда были включены либо лампа А, либо лампа Б. На рисунках 1, 7, 8,9б показан путь одного и того же рабочего, когда была включена лампа А, а на рисунках 2—6, 9а — путь того же муравья, когда он двигался от кормушки к входу при свете лампы Б. г (на всех рисунках) — точки, из которых рабочие замечали отверстие входа в гнездо. На рисунке 10 изображен путь другого рабочего. В этом опыте переключение света производилось в тот момент, когда муравей находился в точке, отмеченной крестом: а — 2-й рейс после переключения (самый продолжительный); б — 10-й рейс после переключения; в — первый рейс после того, как перестали переключать свет
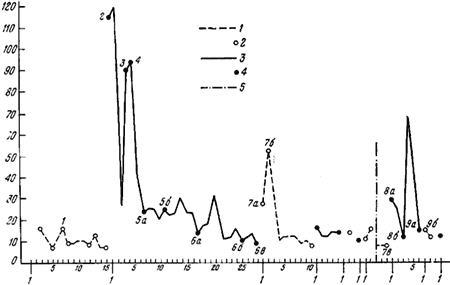
Рис. 83. Реакция рабочих
На горизонтальной оси — порядок опытов, на вертикальной — продолжительность рейсов в секундах. На этом рисунке изображены результаты того же опыта, что и на рис.82/1—9/; цифры на графиках соответствуют номерам этого рисунка; 1,2 — включена только лампа А; 3, 4 — дорогу от кормушки к гнезду муравей находит при свете лампы Б; 5 — перерыв на 1 час. 20 мин. (все время была включена лампа А)
 Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant
Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant(от греч. entoma насекомые и ...логия), раздел зоологии, изучающий насекомых. Основные разделы энтомологии: общая, медицинская, ветеринарная, сельскохозяйственная, лесная.
 Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелец
Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелец Myrmica rubra (Linnaeus, 1758) — рыжая мирмика
Myrmica rubra (Linnaeus, 1758) — рыжая мирмиканадсемейство летающих насекомых отряда перепончатокрылых, родственное осам и муравьям. Существует около 20 тысяч видов пчёл. Распространены на всех континентах, кроме Антарктиды. Пчёлы питаются нектаром и пыльцой, используя нектар главным образом в качестве источника энергии, а пыльцу для получения белков и других питательных веществ.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
 Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравейчасть гнезда рыжих лесных муравьев, занимаемая одной колонной.






