






У муравьев конкуренция между видами, занимающими сходные экологические ниши, может протекать двумя способами. У видов средних размеров, не образующих многочисленных семей, наблюдаются устойчивые сообщества, изученные Понтиным (Pontin, 1961, 1963) для ![]() Lasius niger,
Lasius niger, ![]() L. flavus и Качмареком (Kaczmarek, 1953) для
L. flavus и Качмареком (Kaczmarek, 1953) для ![]() Lasius niger,
Lasius niger, ![]() Myrmica rubra, M. ruginodis, M. scabrinodis, M. schencki, M. rugulosa,
Myrmica rubra, M. ruginodis, M. scabrinodis, M. schencki, M. rugulosa, ![]() Formica fusca. Кормовые участки гнезд всех видов, входящих в такое сообщество, перекрываются. Основным стабилизирующим фактором в этом случае является наличие пищи. Как показал Понтин, количество крылатых половых особей, развивающихся в гнезде, прямо пропорционально площади кормового участка (а следовательно, количеству пищи).
Formica fusca. Кормовые участки гнезд всех видов, входящих в такое сообщество, перекрываются. Основным стабилизирующим фактором в этом случае является наличие пищи. Как показал Понтин, количество крылатых половых особей, развивающихся в гнезде, прямо пропорционально площади кормового участка (а следовательно, количеству пищи).
Рис. 74. Взаимное расположение кормовых участков разных видов Formica (изображены те же участки, что и на рис:76, рис:77)
1—3 — границы кормовых участков: 1 —

Рис. 75. План территорий F. pratensis (pr), F. polyctena (ро) и
1 — границы территорий; 2 — изменение границ после выкладывания сахара (+); 3 — тропы; 4 — границы мелких растительных формаций; I — поляна, сомкнутость травяного покрова 0,7; II — участок леса из дуба и сосны 40 лет, сомкнутость крон 0,7; горизонтальная штриховка — кормовые участки
Показательны опыты, поставленные этим автором. Он затравливал отдельные гнезда ![]() L. niger и наблюдал, какое влияние это окажет на продукцию молодых самок в соседних гнездах
L. niger и наблюдал, какое влияние это окажет на продукцию молодых самок в соседних гнездах ![]() L. niger и
L. niger и ![]() L. flavus. В одном опыте соседние с затравленным гнезда
L. flavus. В одном опыте соседние с затравленным гнезда ![]() L. niger увеличили продукцию самок, что в свою очередь вызвало снижение количества самок в окружающих их гнездах
L. niger увеличили продукцию самок, что в свою очередь вызвало снижение количества самок в окружающих их гнездах ![]() L. flavus. В другом случае, сравнивая численность самок обоих видов (весовое соотношение niger : flavus = 1,47:1), он высчитал, что после затравливания гнезда
L. flavus. В другом случае, сравнивая численность самок обоих видов (весовое соотношение niger : flavus = 1,47:1), он высчитал, что после затравливания гнезда ![]() L. niger из находящегося рядом гнезда
L. niger из находящегося рядом гнезда ![]() L. flavus на следующий год должно было вылететь на 300 самок больше. Фактические результаты (286 самок) подтвердили теоретические расчеты. В одном из гнезд
L. flavus на следующий год должно было вылететь на 300 самок больше. Фактические результаты (286 самок) подтвердили теоретические расчеты. В одном из гнезд ![]() L. niger развивалось 48 самок в 1956 г., а в 1957 г. 107. В результате в соседнем гнезде
L. niger развивалось 48 самок в 1956 г., а в 1957 г. 107. В результате в соседнем гнезде ![]() L. flavus в 1957 г. было на 107 самок меньше (теоретическая цифра — около 90).
L. flavus в 1957 г. было на 107 самок меньше (теоретическая цифра — около 90).
При более или менее постоянных условиях обитания сохраняется стабильность сообщества. Но по мере изменения среды состав сообщества меняется, и одни доминирующие виды сменяются другими. Такими факторами могут быть высота и густота травяного покрова (Pontin, 1963), сомкнутость крон деревьев или наличие крупных видов, таких, как рыжие лесные муравьи (Kaczmarek, 1953).
Из Formica наличие такого типа регуляции показано пока только для ![]() F. fusca (Kaczmarek, 1953). По нашим косвенным наблюдениям (отсутствие охраняемых территорий), так же ведут себя
F. fusca (Kaczmarek, 1953). По нашим косвенным наблюдениям (отсутствие охраняемых территорий), так же ведут себя ![]() F. cunicularia и
F. cunicularia и ![]() F. sanguinea (рис:74).
F. sanguinea (рис:74).
Другой тип межвидовых отношений наблюдается у крупных видов, образующих большие семьи. Эти виды имеют систему охраняемых территорий. Элтон (1934) определяет территорию как систему, регулирующую численность у животных, не имеющих врагов или сравнительно гарантированных от них /У муравьев как единица конкуренции выступает не особь, а семья, которая у Formica s. str. действительно практически не имеет врагов/. В пределы территории не допускаются особи конкурирующих семей другого или того же вида. Система территорий хорошо изучена у птиц и хищных млекопитающих.
У муравьев система территорий, в том смысле, как мы ее сейчас понимаем, впервые была описана Элтоном (С. Elton, 1932) для ![]() F. rufa (s. l.). Мы обнаружили территории у следующих видов:
F. rufa (s. l.). Мы обнаружили территории у следующих видов: ![]() F. rufa, F. polyctena, F. aquilonia, F. pratensis,
F. rufa, F. polyctena, F. aquilonia, F. pratensis, ![]() F. truncorum, F. exsecta, F. foreli, F. forsslundi и F. cinerea (s. str. и imitans) (Длусский, 1965a). Территория у муравьев, как и у других животных, по-видимому, совпадает с кормовым участком. Факторы, определяющие размеры кормового участка, мы рассматриваем в главе, посвященной питанию муравьев, а сейчас остановимся только на вопросах, касающихся конкуренции (рис:75).
F. truncorum, F. exsecta, F. foreli, F. forsslundi и F. cinerea (s. str. и imitans) (Длусский, 1965a). Территория у муравьев, как и у других животных, по-видимому, совпадает с кормовым участком. Факторы, определяющие размеры кормового участка, мы рассматриваем в главе, посвященной питанию муравьев, а сейчас остановимся только на вопросах, касающихся конкуренции (рис:75).
У представителей первой группы при наличии более или менее одинаковых условий границы кормового участка не определенны и обычно представляют собой круг, радиусом которого является максимальное расстояние, на которое рабочие удаляются от гнезда. В зависимости от наличия пищи особи могут добывать ее в той или иной части кормового участка (Pickles, 1934, 1936; Kaczmarek, 1953; Pontin, 1961, 1963). У видов же, имеющих охраняемую территорию, границы ее бывают неправильной формы. Эта форма определяется, помимо характера растительности или наличия пищи, воздействием соседних гнезд конкурирующих видов (С. Elton, 1932; Длусский, 1965а). Наиболее сильные гнезда (при отсутствии влияния растительности) лежат более или менее в центре территорий (![]() F. truncorum на рис:77, F. exsecta на рис:78). Если же гнездо слабое, то территория сильно сдвигается, и смежная граница двух видов проходит недалеко от этого гнезда (F. foreli на рис:76,
F. truncorum на рис:77, F. exsecta на рис:78). Если же гнездо слабое, то территория сильно сдвигается, и смежная граница двух видов проходит недалеко от этого гнезда (F. foreli на рис:76, ![]() F. rufa на рис:78).
F. rufa на рис:78).
Разные виды муравьев по-разному приспособлены к добыванию пищи в разных условиях (Forel, 1874). Поэтому зачастую границы территорий двух конкурирующих видов проходят по границам растительных формаций. Так, например, на участке №4 (Приокско-Террасный заповедник) граница ![]() F. rufa и F. pratensis проходила по границе редкого осинника (сомкнутость 0,2—0,3) с обильным травяным покровом (сомкнутость 0,9— 1, высота 50—100 см), где охотились F. pratensis, и березняка с примесью сосны (сомкнутость 0,7) и с редким травяным покровом, где охотились
F. rufa и F. pratensis проходила по границе редкого осинника (сомкнутость 0,2—0,3) с обильным травяным покровом (сомкнутость 0,9— 1, высота 50—100 см), где охотились F. pratensis, и березняка с примесью сосны (сомкнутость 0,7) и с редким травяным покровом, где охотились ![]() F. rufa. В одних случаях эта граница была ближе к гнезду pratensis, в других — к rufa (рис:76). Таким образом, разные виды оказываются более конкурентоспособными в разных растительных формациях.
F. rufa. В одних случаях эта граница была ближе к гнезду pratensis, в других — к rufa (рис:76). Таким образом, разные виды оказываются более конкурентоспособными в разных растительных формациях.
При изучении территорий муравьев мы обнаружили любопытное явление. У некоторых изученных видов (![]() F. rufa, F. polyctena,
F. rufa, F. polyctena, ![]() F. truncorum, F. exsecta) рабочие относят остатки (точнее, один хитин) съеденных насекомых на границу территории и там выбрасывают. При этом рабочий движется прямо, доходит до границы, оставляет ношу и так же по прямой возвращается в гнездо. Это показывает, что муравьи каким-то образом (скорее всего по запаху) чувствуют границу территории. Точно так же рабочий, зашедший на чужую территорию, пройдя несколько сантиметров, поворачивает обратно, даже в том случае, если не встречает, особей другого вида. Так, например, в течение шести дней в начале августа в Воронежском заповеднике мы проводили наблюдения на участке границы длиной около 3 м между гнездами
F. truncorum, F. exsecta) рабочие относят остатки (точнее, один хитин) съеденных насекомых на границу территории и там выбрасывают. При этом рабочий движется прямо, доходит до границы, оставляет ношу и так же по прямой возвращается в гнездо. Это показывает, что муравьи каким-то образом (скорее всего по запаху) чувствуют границу территории. Точно так же рабочий, зашедший на чужую территорию, пройдя несколько сантиметров, поворачивает обратно, даже в том случае, если не встречает, особей другого вида. Так, например, в течение шести дней в начале августа в Воронежском заповеднике мы проводили наблюдения на участке границы длиной около 3 м между гнездами ![]() F. rufa и F. exsecta. В каждый из дней наблюдения мы могли с точностью до 5 см провести линию, за пределы которой муравьи одного вида не переходят. Лишь очень редко (не более 3 раз в течение часа на участке 1 м) отдельные рабочие заходили на расстояние до 10 см и еще реже — (менее 1 раза) на расстояние до 20 см, хотя на этом участке в пограничной полосе шириной 50 см в середине дня постоянно находилось 35—50 рабочих. Исключением являлся случай, о котором будет сказано ниже.
F. rufa и F. exsecta. В каждый из дней наблюдения мы могли с точностью до 5 см провести линию, за пределы которой муравьи одного вида не переходят. Лишь очень редко (не более 3 раз в течение часа на участке 1 м) отдельные рабочие заходили на расстояние до 10 см и еще реже — (менее 1 раза) на расстояние до 20 см, хотя на этом участке в пограничной полосе шириной 50 см в середине дня постоянно находилось 35—50 рабочих. Исключением являлся случай, о котором будет сказано ниже.
Во всех случаях, которые мы наблюдали, на границе территорий двух видов плотность особей выше, чем в тех участках, где территории не граничат с территориями других видов. Так, например, как уже говорилось выше, на границе территорий ![]() F. rufa и F. exsecta на площадке 0,5 м² в середине дня мы насчитывали одновременно от 35 до 50 рабочих. В то же время на северных участках границы гнезда F. exsecta на каждой из 10 площадок такого же размера находилось не более 10 рабочих и лишь в одном случае, когда на краю территории росла рябина, посещаемая этими муравьями из-за тлей, их было много. Когда мы определяли при помощи ловушек распределение особей на территории гнезда
F. rufa и F. exsecta на площадке 0,5 м² в середине дня мы насчитывали одновременно от 35 до 50 рабочих. В то же время на северных участках границы гнезда F. exsecta на каждой из 10 площадок такого же размера находилось не более 10 рабочих и лишь в одном случае, когда на краю территории росла рябина, посещаемая этими муравьями из-за тлей, их было много. Когда мы определяли при помощи ловушек распределение особей на территории гнезда ![]() F. truncorum (участок №6) в Приокско-террасном заповеднике, оказалось, что за сутки в одну ловушку на границе с территориями других видов попадалось в среднем 7,3±1,6 особей (n=7), тогда как на других участках границы эта величина равнялась 3,1±0,7 (n=8).
F. truncorum (участок №6) в Приокско-террасном заповеднике, оказалось, что за сутки в одну ловушку на границе с территориями других видов попадалось в среднем 7,3±1,6 особей (n=7), тогда как на других участках границы эта величина равнялась 3,1±0,7 (n=8).
Биологический смысл этого явления, по-видимому, в том, что муравьи при малейшей опасности могут быстро собраться вместе для отражения «соседей».
Выкладывая на границе двух территорий кусочки сахара (участок №10), мы заставляли рабочих F. exsecta проникать на территорию ![]() F. rufa. В течение 20—30 мин. после того, как «разведчики» обнаружили сахар недалеко от границы на территории
F. rufa. В течение 20—30 мин. после того, как «разведчики» обнаружили сахар недалеко от границы на территории ![]() F. rufa, около 50 рабочих F. exsecta, собравшись вместе, двинулись к нему. При этом особи
F. rufa, около 50 рабочих F. exsecta, собравшись вместе, двинулись к нему. При этом особи ![]() F. rufa, попавшие в эту массу, уничтожались. Когда же F. exsecta достигли сахара, и установилась новая граница, рабочие вновь рассеялись. Тогда в свою очередь собралась группа рабочих
F. rufa, попавшие в эту массу, уничтожались. Когда же F. exsecta достигли сахара, и установилась новая граница, рабочие вновь рассеялись. Тогда в свою очередь собралась группа рабочих ![]() F. rufa и таким же образом двигалась к сахару и уничтожала попадавшихся рабочих F. exsecta. Через 15 мин. была установлена новая граница.
F. rufa и таким же образом двигалась к сахару и уничтожала попадавшихся рабочих F. exsecta. Через 15 мин. была установлена новая граница.
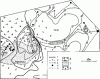
Рис. 76. План территорий F. pratensis,
1 — границы территорий; 2 — изолинии, соединяющие ловушки с одинаковым количеством муравьев в сутки; 3 — тропы к деревьям с тлями; 4 — гнезда; 5 — границы мелких растительных формаций; 6 — высокое разнотравье (сомкнутость травяного покрова 1,0) с редкими осинами; 7 — березовый лес (сомкнутость крон 0,7, сомкнутость травяного покрова 0,7); 8 — березовый лес с примесью сосны (сомкнутость крон 0,7, сомкнутость травяного покрова 0,5)
Рис. 77. План территории
Обозначения те же, что на рис.76
Территория у всех указанных видов Formica охраняется только от тех видов, которые сами имеют территории. Охотничьи участки ![]() F. fusca (рис:74),
F. fusca (рис:74), ![]() F. rufibarbis,
F. rufibarbis, ![]() F. cunicularia, Camponotus vagus, Lasius, Myrmica накладываются на территории этих видов. В одном случае мы наблюдали как
F. cunicularia, Camponotus vagus, Lasius, Myrmica накладываются на территории этих видов. В одном случае мы наблюдали как ![]() F. truncorum и
F. truncorum и ![]() F. fusca посещали колонии тлей на одной и той же сосне, правда, на разных ветках. Поведение
F. fusca посещали колонии тлей на одной и той же сосне, правда, на разных ветках. Поведение ![]() F. fusca также отличается от поведения видов, имеющих охраняемые территории. Рабочие последних, как мы наблюдали неоднократно на границе
F. fusca также отличается от поведения видов, имеющих охраняемые территории. Рабочие последних, как мы наблюдали неоднократно на границе ![]() F. rufa и F. exsecta, встречаясь с рабочими чужих видов, сразу же принимают угрожающую позу, а
F. rufa и F. exsecta, встречаясь с рабочими чужих видов, сразу же принимают угрожающую позу, а ![]() F. fusca — сразу же убегают.
F. fusca — сразу же убегают.
В тех случаях, когда имеется не гнездо, а колония из нескольких гнезд, охраняется территория всей колонии (Мариковский, 1962а). По-видимому, каждое гнездо имеет в пределах этой территории кормовой участок. Но, с другой стороны, несомненно, что данная территория может обеспечить пищей только определенное количество семей. Каким образом происходит эта регуляция — неизвестно.
Насколько границы территории постоянны во времени, пока неясно. Имеются многочисленные данные о том, что рыжие лесные муравьи многие годы используют для разведения тлей одни и те же деревья (Кауцис, 1956). С другой стороны, определение границ территории гнезда F. exsecta, проведенные нами 2 и 8.VIII 1962 г. в Воронежском заповеднике, показало, что границы в эти два дня несколько различались (рис:78).
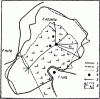
Рис. 78. План территорий F. exsecta и
1 — границы территорий на 8.VIII. 1962 г.; 2 — граница территории F. exsecta на 2.VIII; 3 — граница территорий на 2.VIII на участке, где выкладывался сахар; 4 — гнезда; 5 — тропы к деревьям с тлями; 6 — поляна
Двенадцатичасовые наблюдения на границе гнезд F. exsecta и F. polyctena, расположенных на расстоянии 4 м друг от друга (Воронежский заповедник), показали, что в полдень (температура 25—27°) происходил сдвиг границы в сторону гнезда F. polyctena. После того как температура почвы упала ниже 20°, граница установилась там же, где была утром. Колебания в некоторых местах достигали 20 см. Эту разницу можно считать достоверной, так как визуально всегда можно определить границу с точностью до 5 см.
Установление границ территорий происходит ежегодно после весеннего пробуждения муравьев. В условиях Подмосковья, по нашим наблюдениям, это происходит с середины апреля до конца мая — начала июня. При этом на границах территорий гнезд разных видов постоянно можно наблюдать «бои» между муравьями, ведущие к гибели значительного количества особей обоих муравейников. К середине июля устанавливаются более или менее постоянные границы, и «столкновений» между особями разных видов, как правило, не наблюдается.
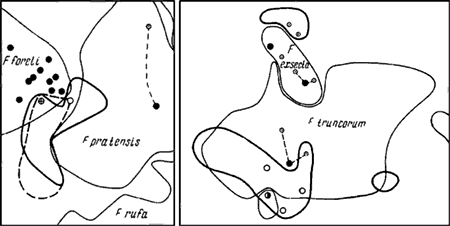
Рис. 74. Взаимное расположение кормовых участков разных видов Formica (изображены те же участки, что и на рис:76, рис:77)
1—3 — границы кормовых участков: 1 —

Рис. 75. План территорий F. pratensis (pr), F. polyctena (ро) и
1 — границы территорий; 2 — изменение границ после выкладывания сахара (+); 3 — тропы; 4 — границы мелких растительных формаций; I — поляна, сомкнутость травяного покрова 0,7; II — участок леса из дуба и сосны 40 лет, сомкнутость крон 0,7; горизонтальная штриховка — кормовые участки
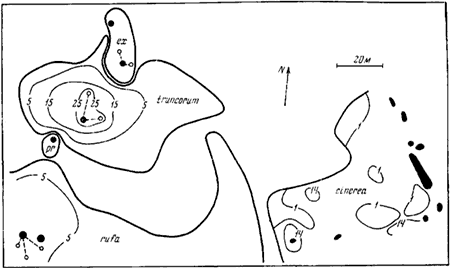
Рис. 77. План территории
Обозначения те же, что на рис:76
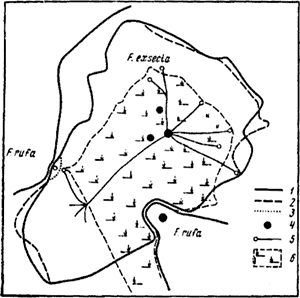
Рис. 78. План территорий F. exsecta и
1 — границы территорий на 8.VIII. 1962 г.; 2 — граница территории F. exsecta на 2.VIII; 3 — граница территорий на 2.VIII на участке, где выкладывался сахар; 4 — гнезда; 5 — тропы к деревьям с тлями; 6 — поляна
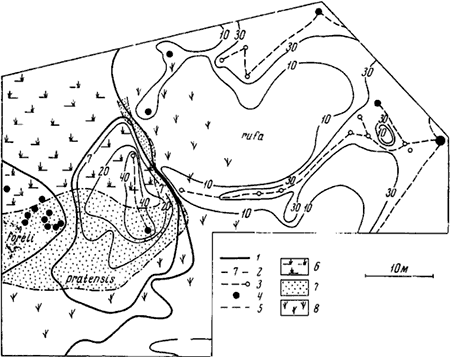
Рис. 76. План территорий F. pratensis,
1 — границы территорий; 2 — изолинии, соединяющие ловушки с одинаковым количеством муравьев в сутки; 3 — тропы к деревьям с тлями; 4 — гнезда; 5 — границы мелких растительных формаций; 6 — высокое разнотравье (сомкнутость травяного покрова 1,0) с редкими осинами; 7 — березовый лес (сомкнутость крон 0,7, сомкнутость травяного покрова 0,7); 8 — березовый лес с примесью сосны (сомкнутость крон 0,7, сомкнутость травяного покрова 0,5)
(от греч. oikos дом, жилище, местопребывание и ...логия), наука об отношениях живых организмов и образуемых ими сообществ между собой и с окружающей средой. Термин «экология» предложен в 1866 Э. Геккелем. Объектами экологии могут быть популяции организмов, виды, сообщества, экосистемы и биосфера в целом.
 Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant
Lasius niger (Linnaeus, 1758) — черный садовый муравей, black ant, garden ant Lasius flavus (Fabricius, 1782) — желтый луговой (садовый, земляной) муравей
Lasius flavus (Fabricius, 1782) — желтый луговой (садовый, земляной) муравей Myrmica rubra (Linnaeus, 1758) — рыжая мирмика
Myrmica rubra (Linnaeus, 1758) — рыжая мирмикаSubspecies: Myrmica scabrinodis ussuriensis, Kuznetsov-Ugamsky
 Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant
Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky antучасток, используемый муравьями для добычи пищи.
 Formica rufibarbis Fabricius, 1793 — краснощекий муравей, краснощекий муравей-минер
Formica rufibarbis Fabricius, 1793 — краснощекий муравей, краснощекий муравей-минер Formica pressilabris Nylander, 1846 — малый тонкоголовый лесной муравей
Formica pressilabris Nylander, 1846 — малый тонкоголовый лесной муравей Formica cunicularia Latreille, 1798 — прыткий степной муравей
Formica cunicularia Latreille, 1798 — прыткий степной муравейучасток, охраняемый семьей (колонной) от посещения чужими особями своего или близких видов.
 Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелец
Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелецосновная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
 Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравей Formica truncorum Fabricius, 1804 — красноголовый муравей
Formica truncorum Fabricius, 1804 — красноголовый муравейполисахарид, образованный остатками аминосахара ацетилглюкозамина. Основной компонент наружного скелета (кутикулы) насекомых, ракообразных и других членистоногих. У грибов заменяет целлюлозу, с которой сходен по химическим и физическим свойствам и биологической роли.
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
Всего 100 видов.
функциональное образование, состоящее из материнской и дочерних семей, поддерживающих регулярные родственные отношения






