






При сравнительном рассмотрении стациального распределения Formica в разных природных зонах можно отметить, что ряд видов подчиняется «принципу смены стаций» (Бей-Биенко, 1920, по Бей-Биенко, 1961). Например, F. lugubris в Карелии обитает в смешанных лесах, а в Московской области — в сырых ельниках, ![]() F. rufa в Карелии — в светлых борах, в Московской области — в смешанных и еловых лесах,
F. rufa в Карелии — в светлых борах, в Московской области — в смешанных и еловых лесах, ![]() F. cunicularia glauca в Воронежском заповеднике — на полянах в сосновых лесах на песках, а в окрестностях Еревана — на заливных лугах,
F. cunicularia glauca в Воронежском заповеднике — на полянах в сосновых лесах на песках, а в окрестностях Еревана — на заливных лугах, ![]() F. fusca в Карелии — только на полянах, в Московской области — на полянах, лугах и под пологом леса, а в Воронежском заповеднике — только под пологом леса. Однако можно найти и исключения из этого правила:
F. fusca в Карелии — только на полянах, в Московской области — на полянах, лугах и под пологом леса, а в Воронежском заповеднике — только под пологом леса. Однако можно найти и исключения из этого правила: ![]() F. truncorum обитает от Карелии до лесостепи преимущественно на полянах сосновых лесов /Об изменении смены стаций у географических подвидов см. главу II (рис:11)/.
F. truncorum обитает от Карелии до лесостепи преимущественно на полянах сосновых лесов /Об изменении смены стаций у географических подвидов см. главу II (рис:11)/.
Каждый вид при продвижении с юга на север или в горах имеет климатические пределы и оптимальную зону обитания. Особенно хорошо это прослеживается в горах (Атанасов, 1953; Длусский, 1962). Паван (Pavan, 1959d) на примере вертикального распределения видов группы ![]() F. rufa в горах Италии наглядно показал, что с продвижением на юг пределы и оптимальная зона обитания поднимаются (рис:72). Сходные данные были получены нами при сравнении вертикальной зональности муравьев Северо-Западного Тянь-Шаня (Таласский Алатау, заповедник «Аксу—Джабаглы»), Юго-Западного Тянь-Шаня (Чаткальский хребет, заповедник «Сары-Челек») и Алайского хребта — Гульча, перевал Талдык, Алайская долина (рис:73).
F. rufa в горах Италии наглядно показал, что с продвижением на юг пределы и оптимальная зона обитания поднимаются (рис:72). Сходные данные были получены нами при сравнении вертикальной зональности муравьев Северо-Западного Тянь-Шаня (Таласский Алатау, заповедник «Аксу—Джабаглы»), Юго-Западного Тянь-Шаня (Чаткальский хребет, заповедник «Сары-Челек») и Алайского хребта — Гульча, перевал Талдык, Алайская долина (рис:73).
В пределах каждой климатической зоны на распределение муравьев влияет ряд факторов, из которых главным является характер растительности (Westhoff, Westhoff de Jonkhere, 1942).
В распределении видов, не способных поддерживать постоянную температуру в гнездах, определяющим фактором является температурный режим почвы в разных стациях. Показательны в этом отношении исследования Э. К. Гринфельда (1939) в заповеднике «Лес на Ворскле» Курской области. На основании данных этого автора нами составлена табл.9, в которой сведены данные, относящиеся к трем видам Serviformica. Как уже отмечалось выше, температурный оптимум ![]() F. cunicularia glauca — 20—32°, F. cinerea imitans — 20—30°, а
F. cunicularia glauca — 20—32°, F. cinerea imitans — 20—30°, а ![]() F. fusca — 27—28°. Поэтому гнезда без земляных холмиков
F. fusca — 27—28°. Поэтому гнезда без земляных холмиков ![]() F. cunicularia glauca наиболее обильны в самом теплом участке, гнезда F. cinerea — в участке с более низким термическим режимом, а
F. cunicularia glauca наиболее обильны в самом теплом участке, гнезда F. cinerea — в участке с более низким термическим режимом, а ![]() F. fusca — в наиболее холодном из этих участков. Постройка холмика позволяет
F. fusca — в наиболее холодном из этих участков. Постройка холмика позволяет ![]() F. cunicularia заселять и стации с более низким режимом температур.
F. cunicularia заселять и стации с более низким режимом температур.
Рис. 72. Оптимальные зоны и верхняя и нижняя границы условий, подходящих для жизни рыжих лесных муравьев (
А — Альпы; Б — Северные Апеннины; В — Южные Апеннины; Г — Сицилия
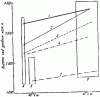
Рис. 73. Изменение вертикальной зональности Formica на разных хребтах гор Средней Азии
А — хр. Таласский Алатау (заповедник « Аксу-Джабаглы»); Б — Чаткальский хребет (Сары-Челекекий заповедник); В — Алайский хребет; 1 — верхняя граница распространения муравьев; 2 — верхняя граница распространения Formica; 3 — граница, ниже которой встречаются более 1 вида Formica; 4 — верхняя граница распространения
Для видов, способных поддерживать в гнезде более или менее постоянную температуру, термический режим почв не играет большой роли (глава VII). Они, как правило, избегают лишь самых крайних условий: сухих полян или лесов с густым подлеском. На распределение этих видов влияет комплекс факторов, из которых определяющими являются возраст и тип.
ТАБЛИЦА 9
Температурный режим почвы различных стаций заповедника «Лес на Ворскле» и распределение Serviformica
(Гринфельд, 1939)
|
Стация
|
Число гнезд на 300 м²
|
Максимальная температура (°С) в 13 час. за лето на глубине (в см)
|
||||||
|
fusca
|
cinerea imitans
|
cunicularia glauca
|
10
|
20
|
30
|
40
|
||
|
без холмика
|
с холмиком
|
|||||||
| Безлесный южный склон |
0
|
0
|
4
|
0
|
35
|
30
|
28
|
27
|
| Пески с мелкой сосной |
0
|
4
|
0
|
0
|
32
|
30
|
26
|
26
|
| Пески с крупной сосной |
1
|
35
|
0
|
0
|
26
|
25
|
22
|
24
|
| Прогалины в дубовом лесу |
1
|
0
|
2
|
10
|
22
|
20
|
20
|
20
|
| Молодой дубовый лес |
2
|
0
|
0
|
1
|
20
|
19
|
18
|
17
|
| Высокоствольный дубовый лес |
1
|
0
|
0
|
0
|
20
|
—
|
19
|
—
|
ТАБЛИЦА 10
Изменение видового состава и сравнительная эффективность* муравьев в разных типах и возрастных группах лесов Европейской части СССР
|
Тип леса
|
Район наблюдений (область)
|
Молодняки
|
Взрослые леса
|
||
|
вид
|
максимальная площадь территорий, га
|
вид
|
максимальная площадь территорий, га
|
||
| Сухие боры | Воронежская, Белгородская |
cinerea |
Более 10***
|
rufa pratensis cinerea |
0,05
0,05 0,05 |
| Московская, Владимирская |
cinerea truncorum |
3***
0,25 |
polyctena |
0,5
|
|
| Свежие боры | Московская, Владимирская |
||||
| Воронежская | exsecta pratensis cinerea** |
0,3
0,15 0,1 |
rufa polyctena |
0,25
0,5 |
|
| Еловые и смешанные насаждения на свежих и влажных почвах | Московская, Владимирская, Воронежская |
exsecta truncorum pratensis** |
0,3
0,5 0,4 |
rufa polyctena aquilonia lugubris |
0,8
2—3 1—2 1—2 |
| Сосновые монокультуры на влажных почвах | Московская,
Воронежская |
rufa pratensis rufa |
0,05
0,05 0,05 |
— |
—
|
** Вид в данных условиях встречается крайне редко.
*** размеры территории указаны для колонии.
Смена видов происходит обычно путем постепенного вытеснения одних видов другими. В зависимости от типа леса смена видов происходит по-разному (табл.10).
В разных типах и возрастных группах леса размеры семей муравьев, а, следовательно, и площадь территорий (кормовых участков), т. е. в конечном счете роль в биоценозах, различны. В табл.10 приведены максимальные территории гнезд различных видов в зависимости от типа леса в молодых и старых лесах.
Formica могут выживать, попадая в неподходящие для них условия, однако это отражается на состоянии семей. Так, F. polyctena в условиях 50—70-летних смешанных насаждений образует колонии из десятков крупных гнезд до 3 м в диаметре. Территории гнезд достигают 2—3 га. Перевезенные нами в 1956 г. в 25-летнюю сосновую монокультуру Воронежского заповедника отводки не погибли, но находятся в угнетенном состоянии. За 6 лет, прошедших после переселения, они остались очень маленькими, ни разу не дали крылатых и территории их не превышают 0,1 га. Они до сих пор остаются одиночными. В то же время в 25-летних сосновых монокультурах F. cinerea находит оптимальные условия. Территории охоты ее колоний охватывают в этих насаждениях десятки гектаров.
Каждый вид муравьев имеет определенные оптимальные условия для добывания пищи, у Formica иногда отличающиеся от оптимальных условий гнездования. Так, например, для F. cinerea в Средней полосе Европейской части СССР для гнезд больше всего подходят хорошо прогреваемые, слабо задернованные (сомкнутость травяного покрова 0,5—0,7) участки, а для охоты эти муравьи предпочитают густые (сомкнутость крон 0,8—1) молодые сосняки, как правило, мертвопокровные. Такие же условия предпочитают для охоты F. polyctena, хотя для гнездования этого вида они совершенно непригодны. Именно этим и объясняется хорошо известный факт, что муравьи чаще всего делают гнезда на границах двух стаций: опушки, края полян, дорог, просек (рис:34). У рыжих лесных муравьев, и особенно у лугового муравья (F. pratensis), от гнезд часто отходят длинные дорожки, идущие в подходящий кормовой участок, с которых фуражиры почти не сходят (рис:75, территория F. pratensis.). Поэтому при неравномерности растительности вокруг гнезда и возможности выбора, границы кормовых участков совпадают с границами изменения растительности.
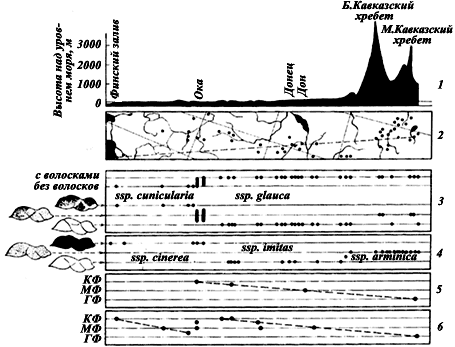
Рис. 11. Географическая изменчивость
1 — профиль рельефа на линии от Еревана до Ленинграда (пунктирная линия на рисунке 2); по вертикали — высота над уровнем моря (в м); 2 — карта полосы шириной 3,5°, проходящей с северо-северо-запада на юго-юго-восток через Европейскую часть СССР и Кавказ, в которой изучалась географическая изменчивость. Точками обозначены места, из которых имеется материал. Эта карта является масштабом для горизонтальной оси всех остальных рисунков; 3 — географическая изменчивость хетотаксии и окраски
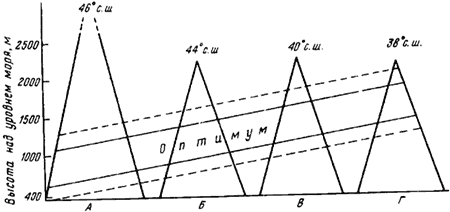
Рис. 72. Оптимальные зоны и верхняя и нижняя границы условий, подходящих для жизни рыжих лесных муравьев (
А — Альпы; Б — Северные Апеннины; В — Южные Апеннины; Г — Сицилия
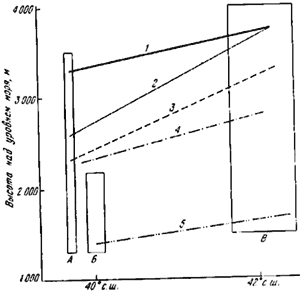
Рис. 73. Изменение вертикальной зональности Formica на разных хребтах гор Средней Азии
А — хр. Таласский Алатау (заповедник « Аксу-Джабаглы»); Б — Чаткальский хребет (Сары-Челекекий заповедник); В — Алайский хребет; 1 — верхняя граница распространения муравьев; 2 — верхняя граница распространения Formica; 3 — граница, ниже которой встречаются более 1 вида Formica; 4 — верхняя граница распространения
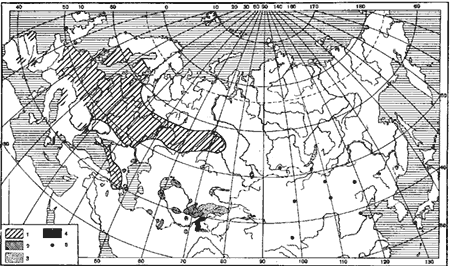
Рис. 34. Географическое распространение
1 —

Рис. 75. План территорий F. pratensis (pr), F. polyctena (ро) и
1 — границы территорий; 2 — изменение границ после выкладывания сахара (+); 3 — тропы; 4 — границы мелких растительных формаций; I — поляна, сомкнутость травяного покрова 0,7; II — участок леса из дуба и сосны 40 лет, сомкнутость крон 0,7; горизонтальная штриховка — кормовые участки
 Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравей Formica cunicularia Latreille, 1798 — прыткий степной муравей
Formica cunicularia Latreille, 1798 — прыткий степной муравей Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant
Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant Formica truncorum Fabricius, 1804 — красноголовый муравей
Formica truncorum Fabricius, 1804 — красноголовый муравейУстарел, исключен.
функциональное образование, состоящее из материнской и дочерних семей, поддерживающих регулярные родственные отношения
участок, используемый муравьями для добычи пищи.
эволюционно сложившаяся, пространственно ограниченная, длительно самоподдерживающаяся, однородная экологическая система, в которой функционально взаимосвязаны живые организмы и окружающая их абиотическая среда.
дочерний муравейник: гнездо, в котором обитает дочерняя семья (в колонии)
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).
размещение (и тип) волосков на теле муравья, отличается большим постоянством и поэтому играет большую роль при определении вида.
 Tetramorium caespitum (Linnaeus, 1758) — дерновый муравей, pavement ant
Tetramorium caespitum (Linnaeus, 1758) — дерновый муравей, pavement antСтарое название: T. indocile
 Formica rufibarbis Fabricius, 1793 — краснощекий муравей, краснощекий муравей-минер
Formica rufibarbis Fabricius, 1793 — краснощекий муравей, краснощекий муравей-минергигрофил






