






Попытаемся теперь на основании весьма ограниченных имеющихся данных сделать некоторые выводы об истории фауны Formica Советского Союза. Как в большинстве построений такого рода, в них будет немало сомнительных предположений. Однако это необходимо сделать для лучшего понимания современной фауны.
Первые Formica появились, по-видимому, в верхнем мезозое в лесах Арктической области (Wheeler, 1913). Это были небольшие муравьи, приблизительно таких же размеров, как современные ![]() F. fusca, у которых рабочие были, вероятно, больше всего похожи на янтарную F. proformicoides. Самки их были сравнительно крупных размеров, поскольку основывали новые семьи самостоятельно.
F. fusca, у которых рабочие были, вероятно, больше всего похожи на янтарную F. proformicoides. Самки их были сравнительно крупных размеров, поскольку основывали новые семьи самостоятельно.
С начала третичного периода в хвойных северных лесах Палеарктики, наряду с родами, встречающимися в настоящее время лишь в тропиках, начала формироваться своеобразная фауна Formica, представленная в балтийском янтаре F. clymene и F. phaethusa, F. antiqua и F. baltica, F. horrida и F. strangulata и не имеющая аналогов в Неарктике. В настоящее время остатками этой древней палеарктической фауны являются, очевидно, F. gagates, F. uralensis, F. sentschuensis и Coptoformica. В период, предшествующий образованию балтийского янтаря, возможно во время эоценового соединения Старого и Нового Света, проникли некоторые виды из Неарктики, где к тому времени сформировалась своя фауна. От этих видов произошла F. flori и группы ![]() F. fusca,
F. fusca, ![]() F. rufibarbis, а также и палеарктические Formica s. str. В отношении Serviformica это предположение базируется на том факте, что неарктические Serviformica, близкие к
F. rufibarbis, а также и палеарктические Formica s. str. В отношении Serviformica это предположение базируется на том факте, что неарктические Serviformica, близкие к ![]() F. fusca, не распадаются на две группы, как это наблюдается в Палеарктике, однако есть и обычная
F. fusca, не распадаются на две группы, как это наблюдается в Палеарктике, однако есть и обычная ![]() F. fusca и виды, близкие к
F. fusca и виды, близкие к ![]() F. rufibarbis, так что можно предположить, что эта дивергенция произошла после того, как предки их были разделены Атлантическим океаном.
F. rufibarbis, так что можно предположить, что эта дивергенция произошла после того, как предки их были разделены Атлантическим океаном.
По-видимому, в нижнем олигоцене уже сформировались подроды и основные группы видов, но Neoformica и группа F. neogagates так и обитали на западе Неарктики и не могли проникнуть в Палеарктику, а Coptoformica обитали в Палеазии, не соединявшейся в то время с Фенноскандией, располагавшейся на месте современной Северной Европы. Образование Raptiformica приходится относить на еще более древнее время, если не считать сходство F. sentschuensis с группой ![]() F. sanguinea конвергентным.
F. sanguinea конвергентным.
В течение третичного периода происходило формирование современной фауны Formica. В начале миоцена на месте нынешней Палеарктики уже вместо раннетретичных лесов были две лесные зоны: Полтавская (тропическая, вечнозеленая) и Тургайская (листопадная, преимущественно занятая широколиственными лесами) (Криштофович, 1946). Граница этих зон проходила в палеогене через нынешние Южную Англию, Башкирию и Северный Казахстан (Марков, 1951). По-видимому, на севере и в горах Тургайской области и происходило формирование палеарктических видов рода Formica.
К плиоцену сформировалась фауна Formica, почти не отличающаяся от современной, и в течение четвертичного периода, вероятно, произошло лишь обособление нескольких видов, вызванное изоляцией популяций: F. mesasiatica в горах Средней Азии от бореальной F. exsecta, F. forsslundi в Европе и F. brunneonitida в Восточной Азии, F. decipiens в Юго-Западной Европе и F. japonica на Дальнем Востоке. Все виды этих пар очень сходны между собой. Разделение F. gagatoides и F. kozlovi, также очень сходных видов, представляется мне следующим образом. Предки этих видов обитали в горах Центральной Азии. Во время одного из ледниковых периодов, ввиду похолодания, они спустились с гор, а когда наступило потепление климата, одни популяции ушли на север, вслед за ледниками, и образовали F. gagatoides, а другие вновь поднялись в горы (F. kozlovi). В Европе F. gagatoides, так же как ![]() F. truncorum, появилась сравнительно недавно, поскольку эти виды обычны на севере континентальной Европы, но отсутствуют на Британских островах.
F. truncorum, появилась сравнительно недавно, поскольку эти виды обычны на севере континентальной Европы, но отсутствуют на Британских островах.
![]() F. sanguinea появилась в Палеарктике в исторически недавние сроки, на что указывает большое сходство этого вида с неарктической F. subnuda Em. Как шло расселение этого вида — не ясно, поскольку он распространен по всей бореальной Палеарктике. Мнение Эмери (Emery, 1920), что это тундрово-степной (?) вид, расселявшийся с запада на восток, на мой взгляд, безосновательно.
F. sanguinea появилась в Палеарктике в исторически недавние сроки, на что указывает большое сходство этого вида с неарктической F. subnuda Em. Как шло расселение этого вида — не ясно, поскольку он распространен по всей бореальной Палеарктике. Мнение Эмери (Emery, 1920), что это тундрово-степной (?) вид, расселявшийся с запада на восток, на мой взгляд, безосновательно.
Гипотетические схемы эволюции Formica, основанные главным образом на морфологических данных, изображены на рис:55, рис:56, рис:57.
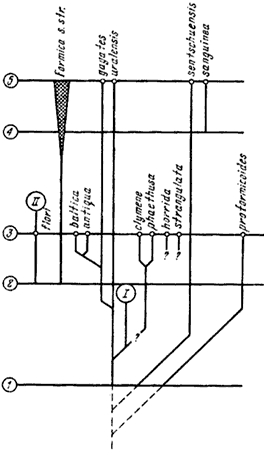
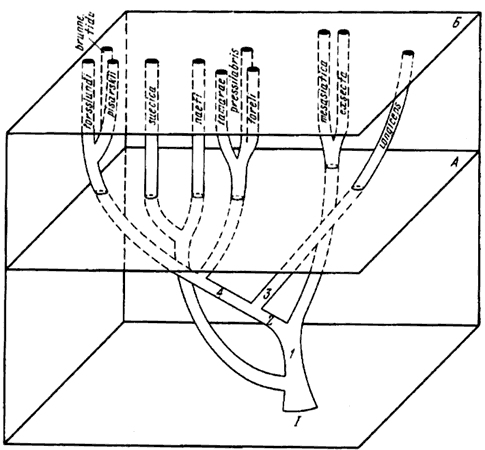
Рис. 56. Гипотетическая схема эволюции Coptoformica
I — см. рис.55; А — граница третичного и четвертичного периодов; Б — современность; 1 — выемка на затылочном крае головы углубляется, затылочные углы становятся приостренными; 2 — уменьшаются размеры самок, укорачиваются нижнечелюстные щупики; 3 — голова удлиняется еще больше; 4 — самки дальше уменьшаются в размерах и почти достигают размеров рабочих, нижнечелюстные щупики укорачиваются дальше

Рис. 57. Гипотетическая схема эволюции групп
Всего 292 вида.
 Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant
Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky antосновная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
Устарел, исключен.
 Formica rufibarbis Fabricius, 1793 — краснощекий муравей, краснощекий муравей-минер
Formica rufibarbis Fabricius, 1793 — краснощекий муравей, краснощекий муравей-минерУстарел, исключен.
Устарел, исключен.
 Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелец
Formica sanguinea Latreille 1798 — кровавый муравей-рабовладелецсовокупность особей одного вида, длительно занимающая определенное пространство и воспроизводящая себя в течение большого числа поколений. В современной биологии популяция рассматривается как элементарная единица процесса эволюции, способная реагировать на изменения среды перестройкой своего генофонда.
 Formica truncorum Fabricius, 1804 — красноголовый муравей
Formica truncorum Fabricius, 1804 — красноголовый муравейнеобратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.






