






Как уже отмечалось выше (глава I), в семействе Formicidae и особенно в роде Formica было описано огромное число варьететов, и до недавнего времени в систематике использовалась сложная квадринарная номенклатура. В результате этого для неспециалиста определение муравьев было непосильной задачей.
Основной задачей, которая была поставлена при ревизии системы рода Formica, было приведение ее в соответствие с правилами зоологической номенклатуры, изложенными в «Международном кодексе зоологической номенклатуры» (1966), что сделало бы возможным точное определение видов для любого квалифицированного энтомолога.
В этой главе мне хотелось бы изложить основные практические принципы, необходимые для понимания современной классификации рода Formica, не углубляясь в вопросы теории. Согласно современным правилам, существуют три основные таксономические категории низшего порядка: род (genus), вид (species) и географический подвид (subspecies). Кроме этого, существуют промежуточные категории надвидового порядка: подрод (subgenus) и группа видов или надвид.
Основной объективной единицей систематики является вид. Можно полностью согласиться с определением вида, данным Майром и др. (1956): «виды представляют собой группы фактически (или потенциально) скрещивающихся естественных популяций, которые репродуктивно изолированы от других таких групп». Однако в практической систематике, как правило, невозможно определить соответствие двух популяций этим требованиям, и большинство систематиков обычно пользуется правилом «разрыва (хиатуса) признаков»: два вида должны абсолютно отличаться по какому-либо признаку. Однако и это правило применимо далеко не всегда.
Как известно, в природе, наряду с так называемыми «хорошими видами», отличающимися от других целым комплексом признаков (например, F. uralensis, F. picea, F. suecica и т. д.), существуют виды-двойники (например, ![]() F. rufa и F. polyctena) и аллопатрические виды (например,
F. rufa и F. polyctena) и аллопатрические виды (например, ![]() F. cunicularia и
F. cunicularia и ![]() F. rufibarbis, F. lemani и
F. rufibarbis, F. lemani и ![]() F. fusca и т. д.), различить которые очень трудно. Особенно усложняет различие этих видов географическая изменчивость.
F. fusca и т. д.), различить которые очень трудно. Особенно усложняет различие этих видов географическая изменчивость.
Как было показано Вильсоном (Wilson, 1955a) на примере Lasius, в ряде случаев у близких видов при отсутствии конкуренции, т. е. в тех географических районах, где встречается только один из них, наблюдается конвергенция по ряду признаков, которая создает видимость межвидовой гибридизации. Однако в местах, где оба эти вида встречаются вместе, они хорошо различаются. Аналогичную картину я обнаружил, анализируя род Formica. Поэтому, следуя за Вильсоном, можно предложить правило, применимое при анализе близких видов: «две формы являются самостоятельными видами, если они отличаются абсолютными признаками (имеется разрыв) в каждой точке, где они обитают вместе».
Поясним это на примере. Ярроу (Yarrow, 1954c) дал полный анализ отличий ![]() F. cunicularia и
F. cunicularia и ![]() F. rufibarbis для Великобритании, который пригоден для большей части Западной Европы. В Европе эти два вида различаются следующими признаками: грудь
F. rufibarbis для Великобритании, который пригоден для большей части Западной Европы. В Европе эти два вида различаются следующими признаками: грудь ![]() F. rufibarbis почти красная, а у
F. rufibarbis почти красная, а у ![]() F. cunicularia — с большими темными пятнами, на груди и чешуйке у rufibarbis имеются отстоящие волоски, а у cunicularia — отсутствуют. По этим признакам эти виды также прекрасно различаются и на большей части Московской области. Однако, когда эти виды были изучены мною со всей территории СССР, то оказалось, что на юге Европейской части СССР, в Сибири и Средней Азии обитает форма с красной грудью и небольшим количеством отстоящих волосков на груди, т. е. по одному из основных признаков приближающаяся к
F. cunicularia — с большими темными пятнами, на груди и чешуйке у rufibarbis имеются отстоящие волоски, а у cunicularia — отсутствуют. По этим признакам эти виды также прекрасно различаются и на большей части Московской области. Однако, когда эти виды были изучены мною со всей территории СССР, то оказалось, что на юге Европейской части СССР, в Сибири и Средней Азии обитает форма с красной грудью и небольшим количеством отстоящих волосков на груди, т. е. по одному из основных признаков приближающаяся к ![]() F. rufibarbis, а по другому — промежуточная между rufibarbis и cunicularia. И только после того, как были отдельно проанализированы материалы из каждого географического района, оказалось возможным установить, что
F. rufibarbis, а по другому — промежуточная между rufibarbis и cunicularia. И только после того, как были отдельно проанализированы материалы из каждого географического района, оказалось возможным установить, что ![]() F. rufibarbis почти не изменяется с продвижением на юг, а
F. rufibarbis почти не изменяется с продвижением на юг, а ![]() F. cunicularia утрачивает темные пятна на груди и у нее появляются отстоящие волоски на груди и чешуйке. Эта форма,
F. cunicularia утрачивает темные пятна на груди и у нее появляются отстоящие волоски на груди и чешуйке. Эта форма, ![]() F. cunicularia glauca, отличается от
F. cunicularia glauca, отличается от ![]() F. rufibarbis тем, что отстоящие волоски у нее, в отличие от
F. rufibarbis тем, что отстоящие волоски у нее, в отличие от ![]() F. rufibarbis, никогда не заходят на заднюю половину среднеспинки (обычно только на переднеспинке), и на чешуйке отстоящие волоски располагаются в один ряд и направлены вверх, а у
F. rufibarbis, никогда не заходят на заднюю половину среднеспинки (обычно только на переднеспинке), и на чешуйке отстоящие волоски располагаются в один ряд и направлены вверх, а у ![]() F. rufibarbis — в два ряда и направлены косо вперед и назад. Телосложение glauca более плотное, чем у других представителей группы
F. rufibarbis — в два ряда и направлены косо вперед и назад. Телосложение glauca более плотное, чем у других представителей группы ![]() F. rufibarbis, и сходно с Formica s. str. или F. gagates. На Кавказе ее заменяет еще один подвид —
F. rufibarbis, и сходно с Formica s. str. или F. gagates. На Кавказе ее заменяет еще один подвид — ![]() F. cunicularia fuscoides с такой же хетотаксией и телосложением, как и glauca, но с темной грудью.
F. cunicularia fuscoides с такой же хетотаксией и телосложением, как и glauca, но с темной грудью.
Виды-двойники (Майр и др., 1956) обычно не имеют абсолютных морфологических различий, но резко отличаются экологически. Морфологически же их можно отличить только по комплексу признаков, имея серию из нескольких экземпляров. Ряд видов Formica, как это вообще часто характерно для муравьев, отличается только по половым особям, а по рабочим они практически неотличимы, по такие формы не могут трактоваться как виды-двойники. Среди палеарктических видов рода Formica видами-двойниками можно считать ![]() F. rufa, F. polyctena и F. aquilonia, для различения которых нельзя дать ни одного абсолютного признака ни для одной из каст.
F. rufa, F. polyctena и F. aquilonia, для различения которых нельзя дать ни одного абсолютного признака ни для одной из каст.
Что касается третьего номена — подвида, то он относится только к географическим подвидам. Трудно не согласиться с мнением Вильсона и Брауна (Wilson, 1955а), что как в случае географически изолированных популяций, так и в случае клинальной изменчивости широко распространенных видов с непрерывным ареалом, не имеет смысла выделять подвиды. Действительно, популяции, существующие длительное время изолированно, всегда в какой-то степени различаются, так что, строго следуя классическим принципам, пришлось бы каждую популяцию описывать как подвид. Что же касается широко распространенных видов с непрерывным ареалом и клинальной (постепенной) географической изменчивостью, то здесь разделение непрерывного ряда на подвиды зависит от желания автора, что и привело к отказу многих исследователей от этой категории (Кузнецов, 1915, Виноградов, 1937, по К. В. Арнольди, 1939; Wilson, 1955а и др.). Ряд авторов (Алпатов, 1927, цит. по К. В. Арнольди, 1939) высказывали мнение, что подвидами нужно называть крайние типы непрерывного ряда, однако, как справедливо отмечает К. В. Арнольди (1939), «далеко не всегда именно крайние типы были установлены раньше промежуточных», так что применение этого принципа повело бы за собой нарушение правил приоритета.
Однако хорошо известен еще один тип клинальной изменчивости, при котором на большой территории вид более или менее однороден (подвид А), а на другой территории так же однороден (подвид В), но отличается от предыдущего, и лишь в узкой промежуточной зоне имеются промежуточные популяции (Huxley, 1940; Шварц, 1963). С. С. Шварц (1963) называет такую изменчивость географически непоследовательной изменчивостью отдельных признаков, а переходные зоны — зонами стабилизации признаков.

Рис. 11. Географическая изменчивость
1 — профиль рельефа на линии от Еревана до Ленинграда (пунктирная линия на рисунке 2); по вертикали — высота над уровнем моря (в м); 2 — карта полосы шириной 3,5°, проходящей с северо-северо-запада на юго-юго-восток через Европейскую часть СССР и Кавказ, в которой изучалась географическая изменчивость. Точками обозначены места, из которых имеется материал. Эта карта является масштабом для горизонтальной оси всех остальных рисунков; 3 — географическая изменчивость хетотаксии и окраски
Автор в одной из своих работ (Dlussky, 1965) использовал менее удачный термин — «ступенчатая» изменчивость.
Вернемся к примеру с ![]() F. cunicularia. Если взять полосу /узкая полоса взята потому, что граница смены подвидов происходит не строго в широтном направлении и к западу изгибается на юг/ шириной около 3,5°, проходящую приблизительно с севера на юг через Европейскую часть СССР (рис:11/1,2/), и проследить изменчивость
F. cunicularia. Если взять полосу /узкая полоса взята потому, что граница смены подвидов происходит не строго в широтном направлении и к западу изгибается на юг/ шириной около 3,5°, проходящую приблизительно с севера на юг через Европейскую часть СССР (рис:11/1,2/), и проследить изменчивость ![]() F. cunicularia в этой полосе, можно увидеть (рис:11/3/), что в узкой зоне между Тулой и Серпуховом происходит смена подвида
F. cunicularia в этой полосе, можно увидеть (рис:11/3/), что в узкой зоне между Тулой и Серпуховом происходит смена подвида ![]() F. cunicularia s. str. подвидом
F. cunicularia s. str. подвидом ![]() F. cunicularia glauca. Аналогичным образом в горах
F. cunicularia glauca. Аналогичным образом в горах ![]() F. cunicularia glauca замещается
F. cunicularia glauca замещается ![]() F. cunicularia fuscoides. Сходное явление наблюдается и у F. cinerea (рис:11/3,4/) с той разницей, что промежуточных особей между F. cinerea s. str. и F. cinerea imitans не найдено (см. главу VI).
F. cunicularia fuscoides. Сходное явление наблюдается и у F. cinerea (рис:11/3,4/) с той разницей, что промежуточных особей между F. cinerea s. str. и F. cinerea imitans не найдено (см. главу VI).
Только в этом смысле я и применяю в данной работе категорию подвида.
На приспособительное значение подвидов указывают многочисленные параллелизмы в географической изменчивости разных видов (Шварц, 1963 и др.). Одним из примеров такого параллелизма может служить рассмотренная выше географическая изменчивость ![]() F. cunicularia и F. cinerea. Как видно из рис:11/3/ и рис:11/4/, зоны стабилизации признаков у этих видов почти совпадают и направление изменений окраски сходно.
F. cunicularia и F. cinerea. Как видно из рис:11/3/ и рис:11/4/, зоны стабилизации признаков у этих видов почти совпадают и направление изменений окраски сходно.
Причины появления подвидов несколько проясняются при попытке построить диаграмму смены стаций. Принцип смены стаций, выдвинутый известным советским энтомологом Г. Я. Бей-Биенко в 20-х годах, состоит в том, что широко распространенные виды, занимающие на севере ареала ксерофитные формации, на юге занимают гигрофитные, что вполне естественно, так как температурные режимы этих формаций сходны (Бей-Биенко, 1961).
Именно такую картину можно наблюдать, сравнивая экологию одного и того же вида с широким ареалом, но не имеющего четких подвидов из разных географических точек, например, ![]() Tetramorium caespitum (рис:11/5/).
Tetramorium caespitum (рис:11/5/).
Но если построить такую же схему для ![]() F. cunicularia (рис:11/6/ высокогорные точки не брались), можно видеть, что смена стаций наблюдается в пределах каждого из подвидов, однако в зоне стабилизации признаков происходит смещение. В окрестностях Ленинграда
F. cunicularia (рис:11/6/ высокогорные точки не брались), можно видеть, что смена стаций наблюдается в пределах каждого из подвидов, однако в зоне стабилизации признаков происходит смещение. В окрестностях Ленинграда ![]() F. cunicularia обитает на песках (ксерофитные стации), а в Московской области на полянах лиственных и смешанных лесов (мезофитные стации). В Воронежской и Липецкой областях
F. cunicularia обитает на песках (ксерофитные стации), а в Московской области на полянах лиственных и смешанных лесов (мезофитные стации). В Воронежской и Липецкой областях ![]() F. cunicularia glauca также встречается только на задернованных полянах сосняков на песках (ксерофитные стации), а в окрестностях Еревана — на сырых лугах по берегам рек (гигрофитные стации). Сходным образом обитающая в долинах речек Тянь-Шаня F. subpilosa litoralis в оазисах и тугаях на равнине Средней Азии сменяется более светлой F. subpilosa s. str. Таким образом, у ряда Serviformica переход от одного подвида к другому сопровождается резкой сменой стации. К сожалению, данные такого рода пока еще крайне скудны, но они заставляют предположить, что образование подвидов, возможно, является средством вида стабилизировать свою стацию.
F. cunicularia glauca также встречается только на задернованных полянах сосняков на песках (ксерофитные стации), а в окрестностях Еревана — на сырых лугах по берегам рек (гигрофитные стации). Сходным образом обитающая в долинах речек Тянь-Шаня F. subpilosa litoralis в оазисах и тугаях на равнине Средней Азии сменяется более светлой F. subpilosa s. str. Таким образом, у ряда Serviformica переход от одного подвида к другому сопровождается резкой сменой стации. К сожалению, данные такого рода пока еще крайне скудны, но они заставляют предположить, что образование подвидов, возможно, является средством вида стабилизировать свою стацию.
Остальные внутривидовые категории, такие как экологические расы (F. polyctena сосновых монокультур и смешанных лесов, F. pratensis и F. nigricans) или цветовые аберрации (меланисты у ![]() F. rufa, красноголовые F. exsecta), могут описываться, но правила приоритета на них не распространяются.
F. rufa, красноголовые F. exsecta), могут описываться, но правила приоритета на них не распространяются.
Что же касается надвидовых категорий, то они в общем довольно субъективны. Дело в том, что между всеми видами имеются разные степени близости, и границы надвидовых категорий будут зависеть от того, на каком уровне различий было произведено разделение. Например, можно, как это делают в последнее время некоторые западные энтомологи (Парашивеску, Гессвальд и др.), назвать подроды родами. В таком случае старые роды становятся надродами. Суть дела от этого не меняется, но система усложняется.
Другое дело, когда новый подрод выделяется на том основании, что столь же хорошо отличающиеся группы в пределах того же рода уже выделены в подроды. Именно на этом основании Мюллер выделил подрод Coptoformica.
В настоящей работе я придерживаюсь наиболее принятой в европейских странах системы, окончательно сформулированной независимо друг от друга Мюллером и Ломницким (G. Muller, 1923; Lomnicki, 1925). Американские авторы не признают некоторых подродов на том основании, что в Неарктике имеются промежуточные формы (Wheeler, 1913; Creighton, 1950), однако часто это связано с незнанием европейской фауны, как, например, в случае с Coptoformica, которые не имеют ничего общего с североамериканской «группой exsecta» (Длусский, 1964а). Точно так же до недавнего времени (Creighton, 1950) американские мирмекологи считали Proformica подродом Formica на основании близости американских представителей к Formica. Однако, если сравнить представителей неарктических «Proformica» с Formica и настоящими Proformica по комплексу признаков, можно видеть, что они гораздо ближе к представителям рода Formica (а именно подроду Serviformica), нежели к настоящим Proformica. С последними они сходны только в строении члеников жгутика усика (Wilson, Brown, 1955). Настоящие Proformica (тип рода P. nasuta Nyl.) — это, по-видимому, чисто палеарктический род центральноазиатского происхождения.
Вильсон и Браун (Wilson, Brown, 1955) уничтожили и подроды Raptiformica и «Proformica» (группа F. neogagates) на том основании, что представители группы F. obtusopilosa по комплексу признаков близки к группе neogagates, но имеют вырезку на переднем крае наличника, как Raptiformica. Таким образом, современные американские авторы оставляют в настоящее время статус подрода только за Neoformica, именуя подроды «группами видов».
Группа видов (надвид) — «это монофилетическая группа очень близких и в значительной мере или полностью аллопатрических видов» (Майр и др., 1956). Мы используем эту категорию для удобства при описании, так как виды, входящие в одну группу, очень сходны. Специальных названий группы не имеют и называются по основному виду («группа ![]() Formica rufa», «группа Formica exsecta» т. д.).
Formica rufa», «группа Formica exsecta» т. д.).
В то же время американские авторы объединяют в группы явно далекие друг от друга виды, например, ![]() F. fusca и F. subpolita и т. д. Если исходить из определений подродовых категорий Майра и др. (1956), «группы видов» американских авторов следует считать подродами.
F. fusca и F. subpolita и т. д. Если исходить из определений подродовых категорий Майра и др. (1956), «группы видов» американских авторов следует считать подродами.
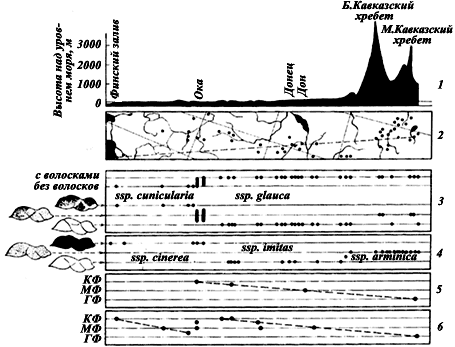
Рис. 11. Географическая изменчивость
1 — профиль рельефа на линии от Еревана до Ленинграда (пунктирная линия на рисунке 2); по вертикали — высота над уровнем моря (в м); 2 — карта полосы шириной 3,5°, проходящей с северо-северо-запада на юго-юго-восток через Европейскую часть СССР и Кавказ, в которой изучалась географическая изменчивость. Точками обозначены места, из которых имеется материал. Эта карта является масштабом для горизонтальной оси всех остальных рисунков; 3 — географическая изменчивость хетотаксии и окраски
МУРАВЬИ - семейство общественных насекомых подотряда жалящих перепончатокрылых. Образуют сложные семьи, включающие несколько каст, выполняющих различные функции (фуражиров, солдат и др.). Длина рабочих муравьев 0,8-30 мм, самки крупнее. Ок. 10 тыс. видов. Более разнообразны и многочисленны в тропиках, северная граница распространения лесотундра. Гнезда в почве, древесине, иногда надземные сооружения (муравейники); существуют и бродячие виды, не имеющие постоянного гнезда. Уничтожают насекомых-вредителей, участвуют в почвообразовании. Во многих странах ряд видов муравьев и их гнезд охраняются.
(от греч. entoma насекомые и ...логия), раздел зоологии, изучающий насекомых. Основные разделы энтомологии: общая, медицинская, ветеринарная, сельскохозяйственная, лесная.
совокупность особей одного вида, длительно занимающая определенное пространство и воспроизводящая себя в течение большого числа поколений. В современной биологии популяция рассматривается как элементарная единица процесса эволюции, способная реагировать на изменения среды перестройкой своего генофонда.
 Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравей Formica cunicularia Latreille, 1798 — прыткий степной муравей
Formica cunicularia Latreille, 1798 — прыткий степной муравей Formica rufibarbis Fabricius, 1793 — краснощекий муравей, краснощекий муравей-минер
Formica rufibarbis Fabricius, 1793 — краснощекий муравей, краснощекий муравей-минер Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky ant
Formica fusca Linnaeus, 1758 — бурый лесной муравей, silky antВсего 100 видов.
часть груди муравья. Состоит из: скутума и скутеллюма. (Извините, информация нуждается в дополнении/уточнении)
часть груди муравья.
размещение (и тип) волосков на теле муравья, отличается большим постоянством и поэтому играет большую роль при определении вида.
(от греч. oikos дом, жилище, местопребывание и ...логия), наука об отношениях живых организмов и образуемых ими сообществ между собой и с окружающей средой. Термин «экология» предложен в 1866 Э. Геккелем. Объектами экологии могут быть популяции организмов, виды, сообщества, экосистемы и биосфера в целом.
у муравьев имеется три основные касты: самцы, самки и рабочие (бесплодные модифицированные самки).
 Tetramorium caespitum (Linnaeus, 1758) — дерновый муравей, pavement ant
Tetramorium caespitum (Linnaeus, 1758) — дерновый муравей, pavement antСтарое название: T. indocile
гигрофил
Устарел, исключен.
Устарел, исключен.
наука изучающая муравьев, часть энтомологии. Мирмекологи - ученые изучающие муравьев.
Всего 24 вида.
Устарел, исключен.






