






20. ![]() Formica rufa Linnaeus, 1761.
Formica rufa Linnaeus, 1761.
Рыжий лесной муравей
![]() F. rufa: Linnaeus, 1761 (non Linnaeus, 1758,= Camponotus herculeanus); F. major Nylander, 1848, Betrem, 1953; F. piniphila Schenck, 1852, Bondroit, 1918; F. truncicola: Forster, 1850 (non Nylander, 1846);
F. rufa: Linnaeus, 1761 (non Linnaeus, 1758,= Camponotus herculeanus); F. major Nylander, 1848, Betrem, 1953; F. piniphila Schenck, 1852, Bondroit, 1918; F. truncicola: Forster, 1850 (non Nylander, 1846); ![]() F. rufa var. major Mayr, 1855;
F. rufa var. major Mayr, 1855; ![]() F. rufa rufa: Forel, 1874 (part.);
F. rufa rufa: Forel, 1874 (part.); ![]() F. rufa rufa var. rufo-pratensis Forel, 1874 (part.);
F. rufa rufa var. rufo-pratensis Forel, 1874 (part.); ![]() F. rufa var. meridionalis Nasonov, 1889 (syn. nov.);
F. rufa var. meridionalis Nasonov, 1889 (syn. nov.); ![]() F. rufa rufa var. meridionalis: Emery, 1909;
F. rufa rufa var. meridionalis: Emery, 1909; ![]() F. rufa rufo-pratensis major: Gosswald, 1941b.
F. rufa rufo-pratensis major: Gosswald, 1941b.
Типовое местонахождение. Европа. Тип (самка) в Британском музее.
Синонимика ![]() F. rufa и F. polyctena. До работ Бетрема (Betrem, 1953), Ланге (Lange, 1960b) и Отто (Otto, 1959с, 1960а) большинство исследователей не отличало эти виды друг от друга. Судя по коллекциям, М. Д. Рузский считал F. polyctena типичной
F. rufa и F. polyctena. До работ Бетрема (Betrem, 1953), Ланге (Lange, 1960b) и Отто (Otto, 1959с, 1960а) большинство исследователей не отличало эти виды друг от друга. Судя по коллекциям, М. Д. Рузский считал F. polyctena типичной ![]() F. rufa, но иногда экземпляры этого вида стоят под этикеткой
F. rufa, но иногда экземпляры этого вида стоят под этикеткой ![]() F. rufa var. rufo-pratensis. Настоящую
F. rufa var. rufo-pratensis. Настоящую ![]() F. rufa этот исследователь определял как
F. rufa этот исследователь определял как ![]() F. rufa var. rufo-pratensis и даже var. truncicolo-pratensis. Описанная В. А. Караваевым из Швеции (1930),
F. rufa var. rufo-pratensis и даже var. truncicolo-pratensis. Описанная В. А. Караваевым из Швеции (1930), ![]() F. rufa var. nuda является полным синонимом F. polyctena (типы в коллекции ЗИ УССР). Вообще же Караваев определял F. polyctena как
F. rufa var. nuda является полным синонимом F. polyctena (типы в коллекции ЗИ УССР). Вообще же Караваев определял F. polyctena как ![]() F. rufa или
F. rufa или ![]() F. rufa var. nuda, а
F. rufa var. nuda, а ![]() F. rufa — как
F. rufa — как ![]() F. rufa,
F. rufa, ![]() F. rufa var. meridionalis и
F. rufa var. meridionalis и ![]() F. rufa var. piniphila.
F. rufa var. piniphila.
Описанная Н. В. Насоновым (1889) из окрестностей Харькова ![]() F. rufa var. meridionalis (типы в ЗМ МГУ) — типичная
F. rufa var. meridionalis (типы в ЗМ МГУ) — типичная ![]() F. rufa. Бледная окраска этих муравьев, возможно, как это считал М. Д. Рузский (1905), является результатом длительного хранения в спирте на свету, но в гнездах
F. rufa. Бледная окраска этих муравьев, возможно, как это считал М. Д. Рузский (1905), является результатом длительного хранения в спирте на свету, но в гнездах ![]() F. rufa иногда встречаются почти такие же светлые экземпляры.
F. rufa иногда встречаются почти такие же светлые экземпляры.

Рис. 45. Географическое распространение
1 — точки нахождения (для Великобритании, Норвегии и Швеции распространение дано вместе для
Согласно синонимике Гессвальда и Г. Шмидта (Gosswald, Schmidt, 1960b), ![]() F. rufa rufo-pratensis minor Gosswald (1941b) (малая раса, «Kleine Rote Waldameise») является: синонимом F. polyctena, а
F. rufa rufo-pratensis minor Gosswald (1941b) (малая раса, «Kleine Rote Waldameise») является: синонимом F. polyctena, а ![]() F. rufa rufa (большая раса, «Grosse Rote Waldameise») —
F. rufa rufa (большая раса, «Grosse Rote Waldameise») — ![]() F. rufa. Но, по-видимому, расы Гессвальда не совпадают точно с этими видами. Средняя раса Гессвальда (
F. rufa. Но, по-видимому, расы Гессвальда не совпадают точно с этими видами. Средняя раса Гессвальда (![]() F. rufa rufo-pratensis major Gosswald, 1941b), очевидно является не самостоятельным видом, как это считал Гессвальд (Gosswald, Schmidt, 1960b), а полигинной формой
F. rufa rufo-pratensis major Gosswald, 1941b), очевидно является не самостоятельным видом, как это считал Гессвальд (Gosswald, Schmidt, 1960b), а полигинной формой ![]() F. rufa.
F. rufa.
В работах Бондруа (Bondroit, 1918) и Бетрема (Betrem, 1953) ![]() F. rufa фигурирует под названием F. piniphila.
F. rufa фигурирует под названием F. piniphila.
Географическое распространение ![]() F. rufa и F. polyctena. В Средней Европе
F. rufa и F. polyctena. В Средней Европе ![]() F. rufa и F. polyctena — наиболее обычные виды рыжих лесных муравьев, но на севере Европы они уступают в численности F. aquilonia, а в Альпах — F. lugubris. В Сибири встречаются реже (
F. rufa и F. polyctena — наиболее обычные виды рыжих лесных муравьев, но на севере Европы они уступают в численности F. aquilonia, а в Альпах — F. lugubris. В Сибири встречаются реже (![]() F. rufa — от Иртыша, F. polyctena — от Оби), а к востоку от Байкала обитает только F. polyctena, да и то встречается редко.
F. rufa — от Иртыша, F. polyctena — от Оби), а к востоку от Байкала обитает только F. polyctena, да и то встречается редко. ![]() F. rufa — единственный вид рыжих лесных муравьев, найденный на Кавказе (в горных лесах), но здесь он редок. В целом F. polyctena распространен дальше на север, чем
F. rufa — единственный вид рыжих лесных муравьев, найденный на Кавказе (в горных лесах), но здесь он редок. В целом F. polyctena распространен дальше на север, чем ![]() F. rufa, а
F. rufa, а ![]() F. rufa встречается южнее, чем F. polyctena (рис:45, рис:46).
F. rufa встречается южнее, чем F. polyctena (рис:45, рис:46).
Географическая изменчивость. ![]() F. rufa отличается большой стабильностью на всем ареале. В Европейской части СССР рабочие южных F. polyctena более волосистые, чем северных, и если на севере Европейской части они очень хорошо отличаются от
F. rufa отличается большой стабильностью на всем ареале. В Европейской части СССР рабочие южных F. polyctena более волосистые, чем северных, и если на севере Европейской части они очень хорошо отличаются от ![]() F. rufa (F. polyctena совершенно без волосков на груди и нижней стороне головы), то в Воронежском заповеднике различить эти виды иной раз бывает затруднительно. Однако в Волгоградской области F. polyctena не отличаются от северных.
F. rufa (F. polyctena совершенно без волосков на груди и нижней стороне головы), то в Воронежском заповеднике различить эти виды иной раз бывает затруднительно. Однако в Волгоградской области F. polyctena не отличаются от северных.
Экология. Как ![]() F. rufa, так и F. polyctena обитают в хвойных, смешанных и лиственных лесах в возрасте свыше 40 лет, но в условиях Средней полосы Европейской части СССР одиночные гнезда
F. rufa, так и F. polyctena обитают в хвойных, смешанных и лиственных лесах в возрасте свыше 40 лет, но в условиях Средней полосы Европейской части СССР одиночные гнезда ![]() F. rufa встречаются в лесах с густым подлеском, а F. polyctena в этих стациях жить не может. Оба вида могут встречаться и на открытых, хорошо прогреваемых полянах и опушках.
F. rufa встречаются в лесах с густым подлеском, а F. polyctena в этих стациях жить не может. Оба вида могут встречаться и на открытых, хорошо прогреваемых полянах и опушках.

Если сравнить количественные соотношения малого и рыжего лесных муравьев в Европейской части СССР, то мы получим следующую картину: в Хибинах встречается только малый муравей; в Архангельской области преимущественно малый муравей, а рыжий лесной — редко; в Московской области оба вида встречаются примерно в одинаковом количестве; в Воронежской области — преимущественно рыжий лесной муравей, хотя и F. polyctena не редок; в Харьковской области встречается только ![]() F. rufa. Исключение представляет пойма Волги, где F. polyctena спускается до Волгограда и здесь доминирует, тогда как
F. rufa. Исключение представляет пойма Волги, где F. polyctena спускается до Волгограда и здесь доминирует, тогда как ![]() F. rufa в Волгоградской области почти не встречается (Щебланов, 1963).
F. rufa в Волгоградской области почти не встречается (Щебланов, 1963).
Форма, материал и размеры гнездовых холмиков варьируют, но в одинаковых условиях гнезда этих видов различаются. Например, в Воронежском заповеднике гнезда F. polyctena всегда имеют вал, а ![]() F. rufa — редко (В Московской области такая закономерность отсутствует). Как правило, F. polyctena строит гнезда из более мелких растительных остатков, нежели
F. rufa — редко (В Московской области такая закономерность отсутствует). Как правило, F. polyctena строит гнезда из более мелких растительных остатков, нежели ![]() F. rufa, и поэтому гнезда этого вида выглядят более аккуратными. Особенно характерно поведение этих видов при искусственном переселении, что мы наблюдали в Воронежском заповеднике совместно с Б. А. Смирновым на переселенных им искусственных отводках. Рабочие F. polyctena в течение короткого времени строят небольшие, но правильные и ровные купола, часто не приурочивая их к пням или стволам деревьев. Рабочие
F. rufa, и поэтому гнезда этого вида выглядят более аккуратными. Особенно характерно поведение этих видов при искусственном переселении, что мы наблюдали в Воронежском заповеднике совместно с Б. А. Смирновым на переселенных им искусственных отводках. Рабочие F. polyctena в течение короткого времени строят небольшие, но правильные и ровные купола, часто не приурочивая их к пням или стволам деревьев. Рабочие ![]() F. rufa строят неправильный рыхлый холмик из веточек, переселяясь к пням, стволам деревьев или штабелям дров.
F. rufa строят неправильный рыхлый холмик из веточек, переселяясь к пням, стволам деревьев или штабелям дров.
![]() F. rufa часто живет одиночными семьями, и большие колонии образует редко, тогда как одиночные гнезда F. polyctena можно встретить лишь как исключение, и в подходящих условиях этот вид образует большие колонии.
F. rufa часто живет одиночными семьями, и большие колонии образует редко, тогда как одиночные гнезда F. polyctena можно встретить лишь как исключение, и в подходящих условиях этот вид образует большие колонии.
Вылет крылатых половых особей происходит на севере Европейской части СССР в конце июня — начале июля, в Московской области в июне и в Воронежской — с середины мая до середины июня. При одинаковых условиях вылет рабочих ![]() F. rufa происходит позже, чем у F. polyctena. Так, например, в окрестностях станции Жаворонки (Московская область) я наблюдал за колониями F. polyctena (группы гнезд ЖА, ЖИ) и
F. rufa происходит позже, чем у F. polyctena. Так, например, в окрестностях станции Жаворонки (Московская область) я наблюдал за колониями F. polyctena (группы гнезд ЖА, ЖИ) и ![]() F. rufa (группы гнезд ЖГ, ЖД, ЖЗ), расположенными в смешанном лесу на легком суглинке и граничащими друг с другом.
F. rufa (группы гнезд ЖГ, ЖД, ЖЗ), расположенными в смешанном лесу на легком суглинке и граничащими друг с другом.
Для изучения фенологии была использована следующая методика: из гнезда бралось 50—100 куколок половых особей и коконы вскрывались. Все куколки разделялись на 4 группы («стадии развития»): 1) предкуколки, 2) белые куколки, 3) желтоватые куколки с темными глазами, 4) почти окрасившиеся куколки. Подсчитывалось количество экземпляров каждой группы и определялся «средний возраст» куколок. Пробы куколок брались с интервалом в несколько дней и фиксировалось изменение «среднего возраста». Линия эта не может быть прямолинейной, так как время прохождения разных «стадий» не одинаково.

Рис. 47. Фенология развития половых особей, в соседних колониях
По горизонтали — дата; по вертикали: L — личинки; 1 — предкуколки; 2 — белые куколки; 3 — желтоватые куколки с темными глазами;4 — почти окрасившиеся куколки; I — имаго; ● —
На рис:47 показана фенология развития куколок в гнездах этих двух колоний. Из графика видно, что к 10.VI в гнездах F. polyctena все половые особи вышли из куколок, а в гнезде ЖА-2 даже вылетели. В то же время 15.VI в гнездах соседней колонии ![]() F. rufa были еще куколки приблизительно 2-й «стадии». В гнездах других колоний
F. rufa были еще куколки приблизительно 2-й «стадии». В гнездах других колоний ![]() F. rufa в том же месте еще были в основном личинки и предкуколки.
F. rufa в том же месте еще были в основном личинки и предкуколки.
Таким образом, можно видеть, что существует фенологическая половая изоляция между соприкасающимися популяциями ![]() F. rufa и F. polyctena.
F. rufa и F. polyctena.
Поскольку F. polyctena и ![]() F. rufa чаще всего встречаются в лесах Европейской части СССР и в Западной Европе, они имеют наибольшее значение для биологической защиты лесов от вредителей. F. polyctena лучше подходит для искусственного расселения, так как чаще образует крупные полигинные семьи и лучше приживается при искусственном расселении. Но и полигинные гнезда
F. rufa чаще всего встречаются в лесах Европейской части СССР и в Западной Европе, они имеют наибольшее значение для биологической защиты лесов от вредителей. F. polyctena лучше подходит для искусственного расселения, так как чаще образует крупные полигинные семьи и лучше приживается при искусственном расселении. Но и полигинные гнезда ![]() F. rufa, особенно из крупных колоний, дают неплохие результаты.
F. rufa, особенно из крупных колоний, дают неплохие результаты.
При искусственном расселении необходимо хорошо различать виды, поскольку, если поселить рядом гнезда разных видов, они начнут враждовать и в результате более слабая семья погибнет, а более сильная будет ослаблена. Вообще всегда при искусственном расселении рыжих лесных муравьев лучше один участок леса заселять отводками, взятыми из одной маточной колонии, поскольку различить эти очень сходные виды часто бывает затруднительно даже при некотором навыке.
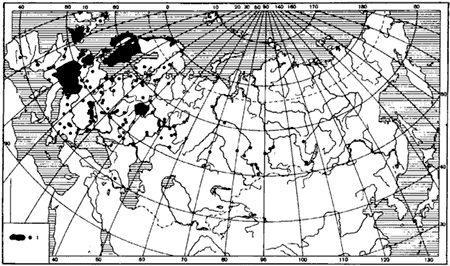
Рис. 45. Географическое распространение
1 — точки нахождения (для Великобритании, Норвегии и Швеции распространение дано вместе для
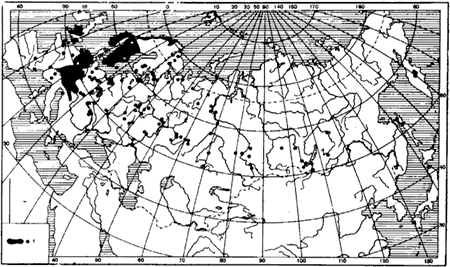
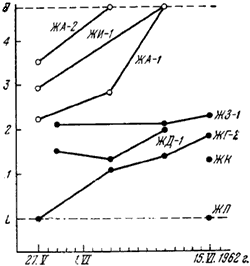
Рис. 47. Фенология развития половых особей, в соседних колониях
По горизонтали — дата; по вертикали: L — личинки; 1 — предкуколки; 2 — белые куколки; 3 — желтоватые куколки с темными глазами;4 — почти окрасившиеся куколки; I — имаго; ● —
 Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравейВсего 965 видов.
наличие в семье нескольких яйцекладущих самок. Смотри: моногиния, олигогиния.
(от греч. oikos дом, жилище, местопребывание и ...логия), наука об отношениях живых организмов и образуемых ими сообществ между собой и с окружающей средой. Термин «экология» предложен в 1866 Э. Геккелем. Объектами экологии могут быть популяции организмов, виды, сообщества, экосистемы и биосфера в целом.
дочерний муравейник: гнездо, в котором обитает дочерняя семья (в колонии)
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
функциональное образование, состоящее из материнской и дочерних семей, поддерживающих регулярные родственные отношения
взрослая особь
совокупность особей одного вида, длительно занимающая определенное пространство и воспроизводящая себя в течение большого числа поколений. В современной биологии популяция рассматривается как элементарная единица процесса эволюции, способная реагировать на изменения среды перестройкой своего генофонда.







