






16. Formica pratensis Retzius, 1783.
Луговой муравей
F. pratensis Retzius, 1783; F. congerens: Forster, 1850 (non Nylander, 1846); ![]() F. rufa var. rufo-pratensis Forel, 1874 (part.);
F. rufa var. rufo-pratensis Forel, 1874 (part.); ![]() F. rufa r. pratensis: Forel, 1874;
F. rufa r. pratensis: Forel, 1874; ![]() F. rufa pratensis var. truncicolo-pratensis Forel, 1874 (part.);
F. rufa pratensis var. truncicolo-pratensis Forel, 1874 (part.); ![]() F. rufa pratensis: Рузский, 1905 (part.);
F. rufa pratensis: Рузский, 1905 (part.); ![]() F. rufa pratensis var. nigricans Emery, 1909;
F. rufa pratensis var. nigricans Emery, 1909; ![]() F. rufa pratensis var. ciliata Рузский, 19156 (non Mayr, 1886) (syn. nov.); F. pratensis var. cordieri Bondroit, 1917;
F. rufa pratensis var. ciliata Рузский, 19156 (non Mayr, 1886) (syn. nov.); F. pratensis var. cordieri Bondroit, 1917; ![]() F. rufa rufo-pratensis minor pratensoides Gosswald, 1951a; F. nigricans: Yarrow, 1954, Gosswald, Schmidt, 1961b etc.; F. cordieri: Gosswald etc., 1961, etc.
F. rufa rufo-pratensis minor pratensoides Gosswald, 1951a; F. nigricans: Yarrow, 1954, Gosswald, Schmidt, 1961b etc.; F. cordieri: Gosswald etc., 1961, etc.
Типовое местонахождение. Европа.
Синонимика. Название F. pratensis Ретциус дал «Fourmi rousse des pres» де Гера, описание которого было настолько неудовлетворительным, что могло быть отнесено к многим видам Formica. Однако все мирмекологи пользовались этим названием до тех пор, пока Ярроу (Yarrow, 1955) не предложил заменить его на F. nigricans Em., поскольку название «F. pratensis Retz.» есть nomen nudum, а первое новое название было предложено Эмери. С этого времени название F. nigricans для этого вида стало появляться на страницах мирмекологической литературы, а после выхода работы Гессвальда и Шмидта (Gosswald, Schmidt, 1960b) полностью вытеснило название F. pratensis. Однако Номенклатурная комиссия II съезда мирмекологов (Вюрцбург, ФРГ) постановила восстановить для этого вида старое название (F. pratensis Retz., 1783), так как оно использовалось в литературе уже более 150 лет и var. nigricans Em. является синонимом var. cordieri Bondr. (Tagungsbericht der Arbeitsgruppe., 1963). F. pratensis, в том объеме, как он рассматривается в настоящей работе, западноевропейские мирмекологи делят на два вида: F. pratensis Retz. (=F. nigricans: Yarrow) и F. nigricans Em. (=F. cordieri Bondr.) (синонимика Номенклатурной комиссии, Tagungsbericht der Arbeitsgruppe..., 1963 на основании изучения типов Эмери и Бондруа). Последняя форма была описана М. Д. Рузским (1915б) под названием ![]() F. rufa pratensis var. ciliata.
F. rufa pratensis var. ciliata.
Рабочие и самцы этих форм почти не отличаются друг от друга, но у F. pratensis самки лишены отстоящих волосков, а у F. nigricans — покрыты обильными отстоящими волосками. Экологических отличий не было обнаружено (Gosswald etc. 1961). На мой взгляд, эти формы являются экологическими расами одного вида, на которые не распространяются правила приоритета. Ниже рассматриваются причины такой точки зрения.
1. Почти на всей территории, где луговой муравей обитает в СССР, во всяком случае во всех точках, где проводились достаточно полные сборы, встречаются обе формы. Анализ литературных данных показывает, что такая же картина наблюдается и в Западной Европе. Лишь в Средней Азии встречается только форма с волосистыми самками.
2. Разделяя эти формы, западноевропейские авторы указывают, что самок с промежуточным характером хетотаксии не встречается (Gosswald etc., 1961). Однако еще Ярроу (Yarrow, 1955) указывал, что такие промежуточные особи встречаются. В коллекциях ЗИН из 44 самок F. pratensis 23 — типичные pratensis, 10 — «nigricans» и 11 — с промежуточным характером хетотаксии. В Воронежском заповеднике в одном из гнезд (B—43) мы обнаружили все три типа самок. Это трудно объяснить гибридизацией, так как гнездо находилось в центре участка соснового леса площадью около 20 га, на котором находилось 21 гнездо формы «nigricans». Во втором гнезде (B—133), из которого была собрана большая серия крылатых самок, часть самок была типа «pratensis», а часть — промежуточные между «nigricans» и «pratensis». Изменчивость самок в некоторых изученных гнездах показана на рис:38.
3. Гессвальдом, Паваном и др. (Gosswald etc., 1961) на основании биометрических исследований итальянских популяций было установлено, что рабочие этих форм отличаются рядом статистических показателей. Наиболее резкие отличия наблюдались при сравнении корреляций длины и ширины головы, что выражается в наклоне линии регрессии. У «pratensis» она наклонена к горизонтальной оси под углом 52—53°, а у «nigricans» — 48—49°. В уравнении линии регрессии (y = a + bx) наклон линии регрессии определяется величиной b, которая является тангенсом угла наклона. Таким образом для итальянских популяций «pratensis» b = 1,28—1,32, а «nigricans» — b = 1,11—1,15.

Для биометрического анализа мною была измерена длина и ширина головы у 104 рабочих F. pratensis, которые были разбиты на четыре группы: A — «pratensis» (Воронежский заповедник), B — промежуточные (Воронежский заповедник), C1 — «nigricans» из Воронежского заповедника, C2 — «nigricans» из Приокско-террасного заповедника. При сопоставлении этих промеров (рис:39) было обнаружено, что в наших условиях нет таких резких различий у рабочих «pratensis» и «nigricans» как в Италии. Хотя угол наклона линии регрессии у «pratensis» больше, чем у «nigricans», его абсолютное значение у популяций «pratensis» из Воронежского заповедника такое же, как у «nigricans» из Италии. Наибольшие достоверные различия найдены для «pratensis» и «nigricans» из Воронежского заповедника (коэффициент b соответственно 1,13±0,005 и 1,06±0,006). Различия между «pratensis» из Воронежского заповедника и «nigricans» из Приокско-террасного заповедника (b = 1,09±0,006) еще меньше. Рабочие из гнезд с промежуточными самками неотличимы от рабочих «nigricans», хотя в гнездах их находятся самки, больше приближающиеся к «pratensis».
4. В условиях Воронежского заповедника наблюдаются различия в экологических требованиях к месту гнездостроения и связанные с этим различия в гнездостроении (см. ниже), однако разница именно такого порядка и должна наблюдаться у экологических рас одного вида.

Рис.39. Корреляция длины и ширины головы рабочих F. pratensis из разных групп гнезд (см. рис.38)
Размеры даны в делениях окуляр-микрометра бинокуляра МБС-1 (10= 0,247 мм);
b = tgα
5. Гессвальд и Шмидт (Gosswald, G. Schmidt, 1960а) наблюдали различия в поведении самцов «pratensis» и «nigricans» при спаривании. Однако сходные этологические различия найдены и у экологических рас F. polyctena (Gosswald, G. Schmidt, 1960b).
Таким образом, видно, что пока нет оснований для разделения этих видов.
Географическое распространение. Западная Европа (в Великобритании только на юге, в Норвегии и Дании не обнаружена); в Европейской части СССР и Западной Сибири северная граница ареала совпадает с северной границей зоны смешанных лесов (за пределами этой границы найдена лишь на Соловецких островах Бирулей), а южная — с южной границей степной зоны; на юге Восточной Сибири до Комсомольска на Амуре, но к востоку от Енисея этот вид встречается очень редко; в горах Средней Азии, но иногда встречается и на равнине, в тугаях рек (рис:40).
Экология. В лесной зоне Европейской части СССР F. pratensis обитает на полянках и просеках сосновых лесов на песчаных и супесчаных почвах, на дюнах (в Прибалтике). В Московской области иногда небольшие гнезда этого вида (r. pratensis) можно встретить и на полянах молодых смешанных лесов на легких суглинках. Разницы в экологии экологических рас в этих условиях мы не обнаружили.

Рис. 40. Географическое распространение F. pratensis
1 — точки нахождения; 2 — южная граница зоны тайги; 3 — южная граница степной зоны; 4 — сомнительные нахождения (F. pratensis или F. lugubris)
В лесостепной зоне (Воронежский заповедник, Галичья гора) раса pratensis лугового муравья обитает преимущественно на остепненных участках и в редких молодых лиственных лесах, а раса nigricans — в сосновых лесах 25—40-летнего возраста, часто под пологом леса.
В лесах луговой муравей обитает в молодых и приспевающих насаждениях и при переходе их в средневозрастные сменяется другими видами. Дело тут не только в конкуренции других видов. Например, в участке сосновой монокультуры на свежей супеси площадью около 40 га в Воронежском заповеднике автором были сделаны полные учеты гнезд Formica s. str. в 1956 и 1962 гг. За это время ни условия освещенности (сомкнутость крон 0,8—0,9), ни характер травянистой растительности (мертвопокровный лес, покрытие менее 0,1) не изменились. Количество гнезд других видов (только ![]() F. rufa) тоже почти не изменилось (было 5 гнезд, стало 6, все по краям квартала, плюс 3 завезенных гнезда F. polyctena с территорией 0,1 га). В то же время в 1956 г., когда насаждения были в возрасте 30 лет, здесь было 32 гнезда F. pratensis, а спустя 6 лет стало 22.
F. rufa) тоже почти не изменилось (было 5 гнезд, стало 6, все по краям квартала, плюс 3 завезенных гнезда F. polyctena с территорией 0,1 га). В то же время в 1956 г., когда насаждения были в возрасте 30 лет, здесь было 32 гнезда F. pratensis, а спустя 6 лет стало 22.
В сибирских лесостепях луговой муравей является эвритопным видом (Рузский, 1946).
М. Д. Рузский, указывая F. pratensis как один из массовых видов степной зоны, однако типичным степняком его не считал, «так как (его) гнезда помещаются всегда на окраинах степи поблизости к кустарникам или опушке леса, куда они неизменно заходят, собирая там обломки сучьев, из которых строят наружные конусы и которыми выполняют внутренность воронок своих гнезд» (стр.78). Однако позднее В. А. Караваев (1935) обнаружил гнезда этого вида в открытой травянистой степи окрестностей Мариуполя (Жданов). Я находил в сходных условиях в окрестностях Изюма (Харьковская область) гнезда лугового муравья, построенные из кусков сухих стеблей конского щавеля и других травянистых растений с деревянистыми стеблями.
На Кавказе и в Крыму обычен и в предгорьях, и в горах, где поднимается до высоты 2500 м.
В горах Средней Азии гнезда F. pratensis встречаются от предгорий (900 м) до высоты 2200 м в степи с кустарником, в ивовых тугаях, луговостепи, розариях, арчевниках и на высокогорных разнотравных лугах. П. И. Мариковский (1962в) изучал биологию этого вида в низовьях р. Или, где он обитает в ивовых и облепиховых тугаях. В среднем течении р. Или этот исследователь, несмотря на тщательные поиски, F. pratensis не обнаружил.
На севере Западной Европы обитает в сосновых лесах на песчаных почвах (Donisthorpe, 1915), в Альпах — от предгорий до высоты 1500 м (Eichhorn, 1964), а на горе Витоша (Болгария) встречается на высоте от 600 до 2000 м, доминируя в смешанных лесах на глинисто-песчаных почвах (Атанасов,1952).
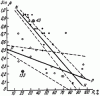
Рис. 41. Зависимость угла наклона пологого ската купола гнезд F. pratensis (β) от количества прямой солнечной радиации (R,%)
N — N, ● — <nigricans>; Р—P, ○ — pratensis; ☺ — гнезда с промежуточными самками. Пунктиром показаны границы изменчивости линии регрессии в пределах ошибки коэффициентов a и b уравнения Sinβ = а + bR
Гнезда лугового муравья обычно имеют плоский купол и широкий земляной вал диаметром до 3 м. Под пологом леса гнезда бывают и с высоким куполом. Как известно, муравьи рода Formica могут менять тепловой режим гнезд, изменяя угол наклона купола к солнцу, т. е. высоту гнезда, и, наоборот, высота гнезд зависит от освещенности. Эта зависимость ближе всего отражается формулой Sinβ = a + bR, где β — угол наклона, пологой стороны купола муравейника, R — количество солнечной энергии падающей на купол (на горизонтальную поверхность), в процентах от максимально возможной для данного места; коэффициент a — положительная величина, b — отрицательная (подробнее см. главу VII).
В условиях Воронежского заповедника мы обнаружили, что у расы nigricans (n = 20) высота гнезд (точнее угол β) больше зависит от освещенности (b = 0,0119±0,0012), при равной освещенности в затененных местах гнезда в среднем выше (а = 1,165±0,035), чем у типичной pratensis (a = 0,528±0,038; b = 0,0039±0,0024; n = 15) (рис:41). Это биологическое приспособление позволяет типичной F. pratensis занимать более освещенные стации, но мешает ей проникать под полог леса, и, наоборот, позволяет расе nigricans существовать в лесах с большой сомкнутостью крон, но делает затруднительным существование этой формы на открытых местах. Возможно, такая зависимость связана с различиями в температурном режиме, необходимом для воспитания молоди.
Гнезда лугового муравья легко отличаются внешне от гнезд рыжих лесных муравьев и красноголового муравья, так как купола их построены из крупных растительных остатков, сцементированных песком, чего никогда не наблюдается у рыжих лесных муравьев (![]() F. rufa). Иногда купол кажется сделанным целиком из песка, среди которого имеются лишь редкие отдельные палочки. Но «внутренний конус», как правило, состоит только из веток. От гнезд к деревьям с колониями тлей идут четкие дорожки, обычно углубленные в земле, а местами даже закрытые сверху. Эти дорожки используются муравьями в течение долгих лет: во Владимирской области я наблюдал, как рабочие одного гнезда лугового муравья пользовались одними и теми же дорожками в течение не менее 15 лет.
F. rufa). Иногда купол кажется сделанным целиком из песка, среди которого имеются лишь редкие отдельные палочки. Но «внутренний конус», как правило, состоит только из веток. От гнезд к деревьям с колониями тлей идут четкие дорожки, обычно углубленные в земле, а местами даже закрытые сверху. Эти дорожки используются муравьями в течение долгих лет: во Владимирской области я наблюдал, как рабочие одного гнезда лугового муравья пользовались одними и теми же дорожками в течение не менее 15 лет.
В питании лугового муравья преобладают мертвые насекомые и падь тлей. Последний вид пищи у него всегда преобладает, и симбиоз с тлями развит у F. pratensis в большей степени, чем у всех других видов Formica s. str. Иногда поедает и живых насекомых, но деятельность его недостаточна для того, чтобы предотвратить вспышки массового размножения хвоегрызущих вредителей. Подробнее питание лугового муравья рассматривается в главе IX.
В условиях Воронежского заповедника рабочие этого вида начинают выходить из гнезд при температуре около 10°, а максимум активности наблюдается при 20°. При повышении температуры до 30° муравьи перестают выходить из гнезда. Популяция F. pratensis, обитающая в низовьях р. Или (Мариковский, 1962в), отличается более высокими температурными требованиями: при 10—12° муравьи вообще перестают двигаться и коченеют.
На большей части ареала в гнездах F. pratensis вылет крылатых половых особей происходит дважды: в мае — июне и в августе — сентябре, чего не наблюдается больше ни у одного из видов Formica. Самки основывают новые семьи паразитически (социальный паразит). Семьи малочисленные по сравнению с рыжими лесными муравьями, и, как правило, моногинные.
М. Д. Рузский (1905) писал, что он находил небольшие колонии этого видав Оренбургских степях. В Европейской части СССР и в Средней Азии я ни разу не находил у этого вида колоний, даже в случае высокой плотности гнезд (Воронежский заповедник); каждое гнездо имеет четко ограниченную территорию и не соединяется дорожками с соседними гнездами (например, см. рис:33/2/).
Для защиты лесов от вредителей луговой муравей практического значения не имеет, так как территория его гнезд в лесах редко превышает 0,15 га, обычно же бывает меньше, и живые насекомые составляют небольшую часть его пищи. Однако другие полезные виды луговой муравей на территории своих гнезд не допускает и мешает им бороться здесь с вредителями.
Одно из перспективных направлений акклиматизации F. polyctena в южных лесах — адаптация самок этого вида в гнездах лугового муравья.
На пастбищах F. pratensis может приносить серьезный вред, так как является промежуточным хозяином ланцетовидной двуустки.
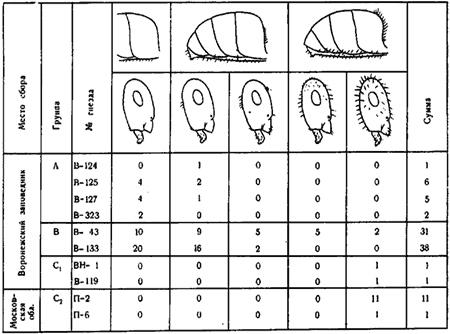
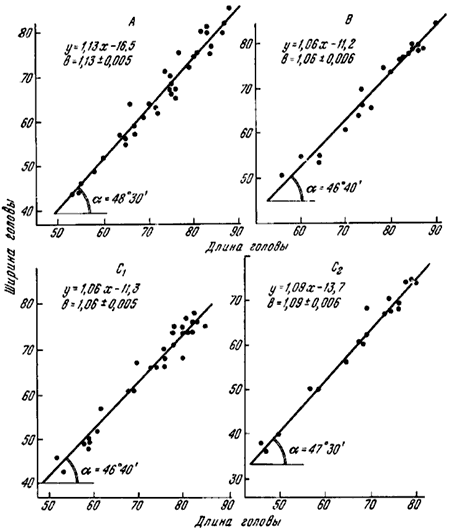
Рис.39. Корреляция длины и ширины головы рабочих F. pratensis из разных групп гнезд (см. рис.38)
Размеры даны в делениях окуляр-микрометра бинокуляра МБС-1 (10= 0,247 мм);
b = tgα
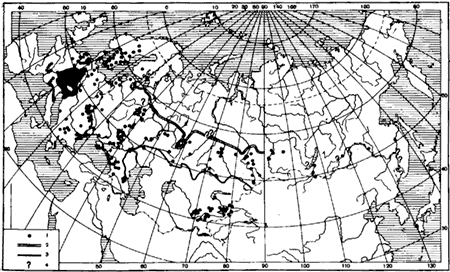
Рис. 40. Географическое распространение F. pratensis
1 — точки нахождения; 2 — южная граница зоны тайги; 3 — южная граница степной зоны; 4 — сомнительные нахождения (F. pratensis или F. lugubris)
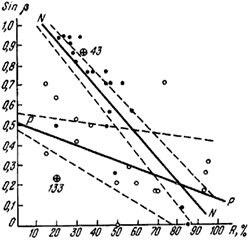
Рис. 41. Зависимость угла наклона пологого ската купола гнезд F. pratensis (β) от количества прямой солнечной радиации (R,%)
N — N, ● — <nigricans>; Р—P, ○ — pratensis; ☺ — гнезда с промежуточными самками. Пунктиром показаны границы изменчивости линии регрессии в пределах ошибки коэффициентов a и b уравнения Sinβ = а + bR
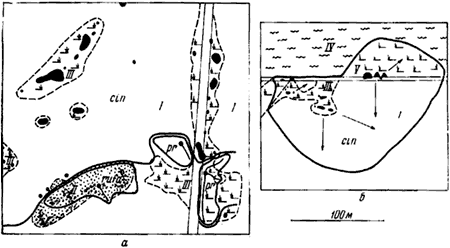
Рис. 33. План территорий двух колоний F. cinerea imitans (Воронежский заповедник)
а — часть колонии В-17 и соседние гнезда других видов; б — колония В-16; I — сосняк мертво-покровный 20—35 лет (сомкнутость крон 0,8—1; 10 С; А1 —А2); II — сосняк мертвопокровный с подростом осины (0,8; 10 С + Ос, Б; А2); III — поляны в этих насаждениях (0,3; 10 С; А1, часто с ягелем); IV — бор-зеленомошник 80—100 лет, распадающийся (0,7; 10 С; A2); V — сосняк злаковый 80—100лет (0,6; 10 С; А1); cin — F. cinerea imitans; pr — F. pratensis; rufa —
 Formica rufa Linnaeus, 1761 — рыжий лесной муравей
Formica rufa Linnaeus, 1761 — рыжий лесной муравейнаука изучающая муравьев, часть энтомологии. Мирмекологи - ученые изучающие муравьев.
(от греч. oikos дом, жилище, местопребывание и ...логия), наука об отношениях живых организмов и образуемых ими сообществ между собой и с окружающей средой. Термин «экология» предложен в 1866 Э. Геккелем. Объектами экологии могут быть популяции организмов, виды, сообщества, экосистемы и биосфера в целом.
размещение (и тип) волосков на теле муравья, отличается большим постоянством и поэтому играет большую роль при определении вида.
совокупность особей одного вида, длительно занимающая определенное пространство и воспроизводящая себя в течение большого числа поколений. В современной биологии популяция рассматривается как элементарная единица процесса эволюции, способная реагировать на изменения среды перестройкой своего генофонда.
(от греч. ethos обычай, нравственный характер и ...логия), биологическая наука, изучающая поведение животных в естественных условиях; уделяет преимущественное внимание анализу генетически обусловленных (наследственных, инстинктивных) компонентов поведения, а также проблемам эволюции поведения.
центральная часть наземного купола у рыжих лесных муравьев. Основное место выделения молоди и обменов между колоннами.
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.
сладкая густая жидкость, выделяемая тлями, червецами и другими насекомыми, питающимися соком растений. Медоносная пчела собирает падь с листьев и перерабатывает в падевый мед.
(от греч. symbíosis — сожительство), в узком смысле под симбиозом понимают такое сожительство особей двух видов, при котором оба партнёра вступают в непосредственное взаимодействие с внешней средой; регуляция отношений с последней осуществляется совместно усилиями, сочетанной деятельностью обоих организмов (ср. Паразитизм). В широком смысле симбиоз охватывает все формы тесного сожительства организмов разных видов, включая и паразитизм, который в этом случае называется антагонистическим симбиозом
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
этап основания семей видов, самки которых неспособны к самостоятельному основанию семьи, на семьях видов-хозяев.
наличие в семье одной яйцекладущей самки. Смотри: полигиния, олигогиния.
функциональное образование, состоящее из материнской и дочерних семей, поддерживающих регулярные родственные отношения
сборное название насекомых подотряда жалящих перепончатокрылых (кроме пчел и муравьев). Длина до 40 мм (шершни). Ок. 23 тыс. видов. Распространены широко. Опылители растений. Личинки некоторых ос паразиты других насекомых.
муравьи ведущие поиск и доставку добычи в муравейник.
Фуражиры активные - муравьи, ведущие самостоятельный поиск добычи и мобилизующие на нее пассивных фуражиров.
Фуражиры пассивные - муравьи, не способные к самостоятельному поиску добычи: мобилизуются на добычу активными фуражирами. Часть пассивных фуражиров имеет закрепленные функции (сборщики пади).






