






ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА МУРАВЬЕВ РОДА MYRMICA(HYMENOPTERA, FORMICIDAE) ЦЕНТРАЛЬНОЙ И ВОСТОЧНОЙ ПАЛЕАРКТИКИ
Приведена определительная таблица муравьев рода Myrmica Центральной и Восточной Палеарктики, включающая 42 вида.
Род Myrmica Latreille — один из наиболее богатых видами родов муравьев Палеарктики, при этом в гумидных районах его представители очень многочисленны практически во всех биогеоценозах. Однако до настоящего времени единой таблицы для определения всех видов на огромных пространствах бывшего СССР не было. Таблицы Рузского (1905) безнадежно устарели.
В последние годы опубликованы современные определительные таблицы мирмик Западной Европы (Agosti, Collingwood, 1987; Seifert, 1988). Имеются определители Myrmica европейской части СССР (Арнольди, 1970; Арнольди, Длусский, 1978), Средней Азии и Южного Казахстана (Арнольди, 1976), Киргизии (Тарбинский, 1976). Туркменистана (Длусский, Союнов, Забелин, 1990) и Дальнего Востока (Купянская, 1990). Однако многие данные, содержащиеся в указанных работах, нуждались в значительном пересмотре. Что касается видов, населяющих территории Сибири и Монголии, то их определение до настоящего времени было практически невозможно.
В приводимую ниже определительную таблицу включены 42 вида мирмик, найденных преимущественно на территории бывшего СССР. Она составлена главным образом на основе признаков рабочих, использование признаков половых особей оговаривается в каждом конкретном случае.
Определение мирмик достаточно сложно и часто требует проведения серии различных промеров. В работе использованы следующие промеры и индексы видов, представленные в таблице:
HL — длина головы от переднего края клипеуса до затылочного края (если он прямой или выпуклый) или до условной линии, соединяющей затылочные углы (если затылочный край вогнутый);
HW — максимальная ширина головы (не включая глаза);
SL — длина скапуса сбоку;
FW — наименьшая ширина лба между лобными валиками;
FLW — наибольшее расстояние между наружными краями лобных лопастей;
TL — длина груди, измеренная в профиль от заднего края метастернальных лопастей до места причленения головы (у рабочих) или до передне-верхнего края проподеума (у самок и самцов);
HI=HL/HW, SI=HL/SL, FI=FLW/FW, FLI=HW/FLW.
|
Вид
|
Рабочие
|
Самцы
|
||||||||||||
|
HL
|
HW
|
SL
|
TL
|
HI
|
SI
|
FI
|
FU
|
HL
|
HW
|
SL
|
IL
|
HI
|
SI
|
|
|
|
1,12—1,26 | 0,91—1,09 | 0,91—1,05 | 1,43—1,81 | 1,15—1,25 | 1,18—1,26 | 1,04—1,13 | 2,12—2,30 | 0,80—1,08 | 0,74—0,97 | 0,70—0,98 | 1,71—1,93 | 1,06—1,13 | 1,10—1,17 |
|
M. ruginodis Nyl.
|
1,09—1,41 | 0,94—1,19 | 0,94—1,18 | 1,62—1,89 | 1,16—1,22 | 1,11—1,25 | 1,09—1,14 | 2,09—2,30 | 0,90—1,13 | 0,84—1,04 | 0,81—1,09 | 1,79—2,13 | 1,05—1,16 | 1,0—1,15 |
|
M. dshungarica Ruzs.
|
0,95—1,23 | 0,78—1,04 | 0,74—0,91 | 1,27—1,65 | 1,18—1,26 | 1,23—1,38 | 1,07—1,17 | 2,07—2,32 | 0,87—1,01 | 0,77—0,85 | 0,77—0,91 | 1,64—1,89 | 1,08—1,20 | 1,03—1,21 |
|
M. kryzhanovskii Arn.
|
1,04—1,16 | 0,85—0,99 | 0,87—0,98 | 1,48—1,61 | 1,16—1,24 | 1,15—1,23 | 1,10—1,11 | 2,18—2,41 |
—
|
—
|
—
|
—
|
—
|
—
|
|
M. juglandeti Arn.
|
1,08—1,18 | o,83—0,97 | 0,81—0,92 | 1,37—1,55 | 1,16—1,25 | 1,16—1,31 | 1,10—1,17 | 2,21—2,67 | 0,76—0,88 | 0,66—0,78 | 0,67—0,85 | 1,53—1,71 | 1,10—1,16 | 1,03—1,16 |
|
M. ferganensis Karaw.
|
0,95—1,02 | 0,77—0,84 | 0,77—0,84 | 1,33—1,48 | 1,20—1,24 | 1,20—1,24 | 1,11—1,13 | 2,22—2,32 |
—
|
—
|
—
|
—
|
—
|
—
|
|
M. arnoldii Dlussky
|
0,92—1,06 | 0,77—0,91 | 0,67—0,74 | 1,23—1,43 | 1,14—1,20 | 1,42—1,50 | 1,21—1,28 | 2,04—2,21 | 0,77—0,78 | 0,73—0,74 | 0,29—0,31 | 1,40—1,57 | 1,04—1,06 | 2,55—2,62 |
|
M. tschekanovskii Radtsch.
|
1,05 | 0,92 | 0,84 | 1,47 | 1,14 | 1,25 | 1,15 | 2,44 | 1,02 | 1,02 | 0,73 | 1,89 | 1,0 | 1,40 |
|
M. kurokii Forel
|
1,33—1,53 | 1,19—1,47 | 1,15—1,26 | 1,92—2,17 | 1,09—1,15 | 1,16—1,27 | 1,00—1,02 | 2,07—2,33 | 1,19—1,22 | 1,08—1,12 | 1,12—1,13 | 2,27—2,30 | 1,09—1,10 | 1,06—1,07 |
|
M. luteola Kupian.
|
1,33—1,51 | 1,34—1,51 | 1,12—1,30 | 1,92—2,33 | 0,98—1,03 | 1,14—1,21 | 1,05—1,08 | 2,40—2,54 | 0,92—0,99 | 0,91—0,99 | 0,39—0,45 | 2,03—2,24 | 1,0—1,02 | 2,13—2,37 |
|
M. schencki Emery
|
1,13—1,29 | 1,01—1,23 | 0,84—1,05 | 1,61—1,99 | 1,0—1,16 | 1,14—1,33 | 1,58—1,94 | 3,61—4,88 | 0,84—0,94 | 0,80—0,92 | 0,31—0,42 | 1,96—2,25 | 1,02—1,06 | 2,03—2,91 |
|
M. lacustris Ruzs.
|
1,15—1,34 | 1,02—1,25 | 0,91—1,09 | 1,72—2,10 | 1,04—1,14 | 1,17—1,29 | 1,35—1,46 | 2,92—3,50 | 0,85—0,94 | 0,83—0,91 | 0,25—0,29 | 2,06—2,28 | 1,02—1,07 | 3,14—3,61 |
|
M. caucasicola Arn.
|
1,19—1,26 | 1,04—1,12 | 0,88—0,98 | 1,61—1,76 | 1,11—1,15 | 1,28—1,35 | 1,47—1,57 | 3,76—4,00 | 1,02—1,08 | 0,97—1,01 | 0,38—0,45 | 2,24—2,35 | 1,06—1,07 | 2,41—2,70 |
|
M. ravasinii Finzi
|
1,29—1,50 | 1,25—1,39 | 1,16—1,26 | 1,96—2,11 | 1,03—1,10 | 1,08—1,18 | 2,13—2,74 | 5,56—7,38 | 0,78—1,04 | 0,97—1,02 | 0,36—0,41 | 2,23—2,34 | 1,01—1,03 | 2,48—2,76 |
|
M. lobicornis Nyl.
|
1,05—1,22 | 0,90—1,16 | 0,83—0,90 | 1,45—1,60 | 1,05—1,17 | 1,20—1,30 | 1,43—1,62 | 3,05—4,10 | 0,89—0,99 | 0,85—0,96 | 0,74—0,87 | 1,77—1,99 | 1,05—1,12 | 1,27—1,45 |
|
M. jessensis Forel
|
0,98—1,18 | 0,87—1,05 | 0,77—0,98 | 1,40—1,68 | 1,10—1,16 | 1,17—1,32 | 1,28—1,42 | 2,65—2,95 | 0,94—1,02 | 0,90—0,98 | 0,62—0,71 | 1,82—1,93 | 1,04—1,06 | 1,43—1,55 |
|
M. kamtschatica Kupiansk.
|
1,04—1,15 | 0,92—1,04 | 0,80—0,90 | 1,51—1,64 | 1,11—1,13 | 1,28—1,34 | 1,22—1,33 | 2,63—2,81 | 0,83—0,94 | 0,78—0,91 | 0,67—0,77 | 1,67—1,89 | 1,02—1,05 | 1,17—1,23 |
|
M. forcipata Karaw.
|
0,95—1,11 | 0,85—0,99 | 0,77—0,90 | 1,41—1,61 | 1,10—1,12 | 1,20—1,25 | 1,27—1,36 | 2,51—2,91 | 0,94—0,97 | 0,91—0,94 | 0,71—0,73 | 1,93—1,96 | 1,03 | 1,31—1,32 |
|
M. pisarskii Radtsch.
|
0,85—1,06 | 0,76—0,97 | 0,69—0,84 | 1,36—1,57 | 1,10—1,16 | 1,24—1,35 | 1,28—1,40 | 2,48—2,75 | 0,81—0,94 | 0,74—0,90 | 0,64—0,69 | 1,54—1,85 | 1,05—1,09 | 1,22—1,36 |
|
M. zojae Radtsch.
|
0,94—1,05 | 0,81—0,94 | 0,78—0,90 | 1,34—1,47 | 1,12—1,19 | 1,17—1,24 | 1,27—1,35 | 2,83—3,10 | 0,78 | 0,76 | 0,64 | 1,62—1,65 | 1,04 | 1,22 |
|
M. kirghisorum Arn.
|
1,09—1,18 | 0,91—1,02 | 0,87—0,93 | 1,42—1,58 | 1,12—1,16 | 1,16—1,29 | 1,24—1,44 | 2,52—2,95 | 0,91—0,98 | 0,78—0,83 | 0,84—0,91 | 1,80—1,96 | 1,08—1,13 | 1,13—1,18 |
|
M. angulinodis Ruzs.
|
0,90—1,13 | 0,77—1,09 | 0,74—0,95 | 1,26—1,64 | 1,04—1,17 | 1,19—1,28 | 1,16—1,23 | 2,39—2,71 | 0,85—0,99 | 0,87—0,97 | 0,69—0,78 | 1,61—1,90 | 0,98—1,06 | 1,21—1,35 |
|
M. saposhnikovi Ruzs.
|
0,95—1,13 | 0,83—1,01 | 0,77—0,91 | 1,34—1,60 | 1,09—1,14 | 1,22—1,28 | 1,19—1,30 | 2,50—2,87 | 0,84—0,88 | 0,81—0,87 | 0,71—0,74 | 1,68—1,76 | 1,0—1,03 | 1,15—1,21 |
|
M. bicolor Kupiansk.
|
1,08—1,16 | 0,90—1,04 | 0,80—0,85 | 1,54—1,61 | 1,11—1,20 | 1,33—1,37 | 1,21—1,26 | 2,48—2,67 | 0,78—0,92 | 0,76—0,88 | 0,57—0,71 | 1,71—1,89 | 1,04—1,07 | 1,29—1,37 |
|
M. kasczenkoi Ruzs.
|
0,95—1,08 | 0,87—0,97 | 0,76—0,83 | 1,36—1,58 | 1,09—1,17 | 1,24—1,33 | 1,18—1,31 | 2,35—2,46 | 0,84—0,85 | 0,76—0,78 | 0,63—0,71 | 1,75—1,83 | 1,07—1,11 | 1,30—1,34 |
|
M. transsibirica Radtsch.
|
1,01—1,13 | 0,85—1,02 | 0,80—0,90 | 1,43—1,64 | 1,10—1,18 | 1,21—1,36 | 1,07—1,15 | 2,28—2,59 | 0,83—0,97 | 0,78—0,95 | 0,60—0,77 | 1,65—1,93 | 1,0—1,08 | 1,16—1,44 |
|
M. sulcinodis Nyl.
|
1,01—1,33 | 0,92—1,19 | 0,85—1,08 | 1,54—1,99 | 1,06—1,14 | 1,18—1,40 | 1,06—1,18 | 2,27—2,66 | 0,98—1,16 | 0,90—0,99 | 0,91—1,02 | 2,02—2,37 | 1,03—1,17 | 1,01—1,22 |
|
M. aspersa Kupian.
|
0,95—1,20 | 0,84—1,12 | 0,83—1,05 | 1,40—1,85 | 1,06—1,18 | 1,11—1,24 | 1,06—1,19 | 2,39—2,71 | 0,83—0,90 | 0,78—0,85 | 0,60—0,73 | 1,86—2,07 | 1,05—1,13 | 1,22—1,45 |
|
M. commarginata Ruzs.
|
1,05—1,09 | 0,88—0,92 | 0,81—0,87 | 1,48—1,54 | 1,18—1,20 | 1,26—1,29 | 1,07—1,11 | 2,25—2,37 |
—
|
—
|
—
|
—
|
—
|
—
|
|
M. scabrinodis Nyl.
|
1,08—1,20 | 0,88—1,09 | 0,80—0,92 | 1,44—1,72 | 1,09—1,21 | 1,24—1,33 | 1,41—1,68 | 2,60—3,04 | 0,78—0,95 | 0,76—0,92 | 0,31—0,41 | 1,68—2,27 | 1,0—1,04 | 2,29—2,95 |
|
M. sabuleti Mein.
|
1,04—1,26 | 0,92—1,16 | 0,84—1,01 | 1,44—1,78 | 1,07—1,15 | 1,22—1,30 | 1,59—1,86 | 3,04—3,50 | 0,84—0,97 | 0,81—0,95 | 0,45—0,56 | 1,79—2,12 | 1,0—1,05 | 1,50—1,97 |
|
M. slovaca Sadil
|
0,97—1,18 | 0,90—1,09 | 0,78—0,92 | 1,37—1,65 | 1,04—1,14 | 1,21—1,31 | 1,70—2,06 | 3,29—3,94 | 0,78—0,84 | 0,78—0,83 | 0,27—0,33 | 1,85—2,02 | 0,98—1,01 | 2,60—2,90 |
|
M. bessarabica Nasonov
|
0,94—1,18 | 0,80—1,04 | 0,76—0,95 | 1,33—1,65 | 1,06—1,19 | 1,15—1,30 | 1,11— 1,41 | 2,19—2,94 | 0,76—0,85 | 0,70—0,83 | 0,29—0,42 | 1,71—1,96 | 1,0—1,10 | 2,03—2,57 |
|
M. stangeana Ruzs.
|
1,04—1,25 | 0,95—1,15 | 0,87—1,01 | 1,54—1,72 | 1,06—1,12 | 1,18—1,28 | 1,31—1,44 | 2,76—3,04 | 0,78—0,80 | 0,76—0,78 | 0,22—0,25 | 1,82—1,86 | 1,02—1,04 | 3,17—3,50 |
|
M. bergi Ruzs.
|
1,04—1,37 | 0,94—1,23 | 0,88—1,13 | 1,54—2,10 | 1,06—1,14 | 1,12—1,24 | 1,06—1,14 | 1,95—2,18 | 0,88—0,99 | 0,80—0,96 | 0,49—0,59 | 1,95—2,18 | 1,04—1,11 | 1,69—1,97 |
|
M. gallienii Bondr.
|
1,05—1,26 | 0,92—1,15 | 0,88—1,05 | 1,44—1,82 | 1,06—1,14 | 1,18—1,26 | 1,05—1,12 | 2,06—2,17 | 0,88—1,09 | 0,83—1,09 | 0,34—0,45 | 1,96—2,31 | 1,0—1,08 | 2,44—2,75 |
|
M. rugulosa Nyl.
|
0,91—1,18 | 0,78—1,05 | 0,70—0,95 | 1,29—1,47 | 1,11—1,20 | 1,23—1,32 | 1,05—1,10 | 1,93—2,06 | 0,76—0,81 | 0,71—0,78 | 0,28—0,35 | 1,65—1,96 | 1,04—1,07 | 2,28—2,70 |
|
M. tenuispina For.
|
0,92—1,19 | 0,74—1,01 | 0,74—0,92 | 1,26—1,62 | 1,17—1,26 | 1,16—1,28 | 1,17—1,36 | 2,25—2,65 | 0,80—0,85 | 0,70—0,77 | 0,29—0,32 | 1,61—2,16 | 1,09—1,14 | 2,52—2,76 |
|
M. orthostyla Arn.
|
1,05—1,12 | 0,88—0,98 | 0,88—0,94 | 1,51—1,61 | 1,14—1,19 | 1,19—1,23 | 1,25—1,30 | 2,33—2,52 | 0,84—0,85 | 0,76—0,77 | 0,28—0,29 | 1,88—1,89 | 1,11 | 2,90—3,0 |
|
M. carinata Kupian.
|
0,99—1,08 | 0,85—0,95 | 0,80—0,90 | 1,44—1,54 | 1,13—1,18 | 1,20—1,25 | 1,06—1,14 | 2,10—2,30 |
—
|
—
|
—
|
—
|
—
|
—
|
|
M. excelsa Kupian.
|
1,11— 1,15 | 0,97—1,04 | 0,83—0,90 | 1,65—1,74 | 1,09—1,13 | 1,28—1,34 | 1,05—1,11 | 2,22—2,39 |
—
|
—
|
—
|
—
|
—
|
—
|
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДА MYRMICA LATREILLE ЦЕНТРАЛЬНОЙ И ВОСТОЧНОЙ ПАЛЕАРКТИКИ (РАБОЧИЕ)
1(4) Членики стебелька рабочих и самок снизу с лопастевидными выступами (рис:3/а,б/). Самцы: антенны 12-члениковые.
2(3) Скапус длиннее (SI=1,25), с уголком на сгибе (рис:1/б/); лопастевидные выступы снизу члеников стебелька развиты слабее (рис:3/б/). Самцы: скапус длинный (SI=1,40). — Красноярский край
M. tschekanovskii Radtschenko, 1994
3(2) Скапус короче (SI=1,42—1,50), без уголка на сгибе (рис:1/а/); лопастевидные выступы снизу члеников стебелька развиты сильнее (рис:3/а/). Самцы: скапус короткий (SI=2,55—2,62). — Южная Сибирь, Северная Монголия
M. arnoldii Dlussky, 1963
4(1) Членики стебелька рабочих и самок снизу без лопастевидных выступов. Самцы: антенны 13-членниковые.
5(80) Боковые края клипеуса не приподняты килевидно, передняя часть усиковой впадины лежит на одном уровне с боковыми краями клипеуса (рис:3/а,и,м,о/; рис:6/а,б/).
6(61) Усиковые ямки окаймлены дуговидными морщинками, либо являющимися продолжением загибающихся наружу лобных валиков, либо причленяющимися к ним (рис:3/е,и,м,о/).
7(8) Проподеум самок без шипов, лишь с тупыми бугорками (рис:3/в/). Половые особи мельче рабочих. Скапус рабочих согнут довольно круто, но без уголка на сгибе (рис:1/к/); голова субквадратная (HI=0,98—1,03); грудь сверху без морщинок, с грубой неправильной петлеобразной скульптурой. Мезопроподеальное вдавление резкое; все тело, ноги и скапус с длинным густым отстоящим опущением. Тело оранжевое или оранжево-бурое. Размеры крупные: HL=1,33—1,51; TL=1,92—2,33 мм. — Приморье, о-в Кунашир
M. luteola Kupianskaya, 1990
8(7) Проподеум самок всегда с явственными шипами (рис:3/г/). Половые особи крупнее рабочих. Набор признаков рабочих иной.
9(22) Скапус согнут слабо и плавно, без уголка на сгибе (рис:1/в—и/). Самцы: скапус длинный, далеко выступает за затылочный край головы (SI=1,0—1,21).
10(11) Крупнее (HL=1,33—1,53; TL=1,92—2,17 мм), скапус согнут круче (рис:1/и/). Мезопроподеальное вдавление слабое, петиолюс без передней цилиндрической части, массивный (рис:3/д/). — Сахалин, Курилы, Япония
M. kurokii Forel, 1907
11(10) Мельче (HL=1,04—1,41; TL=1,92—2,17 мм), скапус согнут слабо (рис:1/в—з/). Мезопроподеальное вдавление резкое, петиолюс с явственной цилиндрической частью, часто длинной. Тело от желтого до красно-бурого.
12(15) Лобные валики сзади загибаются наружу и явственно переходят в резкую морщинку, окаймляющую усиковую впадину (рис:3/е/). Размеры в среднем крупнее (TL=1,43—1,89 мм).
13(14) Узелки стебелька гладкие или со слабой поверхностной скульптурой, без резких морщинок; шипы проподеума короткие (рис:3/ж/). Самцы: скапус и голени с длинными обильными отстоящими волосками (рис:2/в'/). — Транспалеаркт
![]() M. rubra (Linnaeus, 1758)
M. rubra (Linnaeus, 1758)
14(13) Узелки стебелька с резкими морщинками, шипы проподеума длинные (рис:3/з/). Самцы: скапус и голени с короткими редкими отстоящими волосками (рис:2/г'/). — Транспалеаркт
M. ruginodis Nylander, 1846
15(12) Лобные валики прослеживаются по крайней мере до уровня верхнего края глаз; морщинки, окаймляющие усиковые впадины, причленяются к лобным валикам в их верхней трети (рис:3/и,м,о/). В среднем мельче (TL не более 1,65 мм).
16(17) Петиолюс с короткой цилиндрической частью, его передняя поверхность крутая, узелок с горизонтальной или скошенной назад вершинной площадкой (рис:3/д/). Голова с выпуклыми боковыми сторонами. Передний край клипеуса вытянутый, заостренный (рис:3/и/). Самцы: петиолюс с высоким кубическим узелком (рис:3/л/). — Тянь-Шань, Алай
M. dshungarica Ruzsky, 1905
17(16) Петиолюс с длинной цилиндрической частью, его узелок на вершине закругленно-треугольный. Голова с параллельными боковыми сторонами, передний край клипеуса слабо вытянут, закругленный (рис:3/м/). Самцы: петиолюс низкий, явственно ниже постпетиолюса, его узелок закругленно-треугольный (рис:3/м/).
18(19) Скапус с длинными торчащими отстоящими волосками, длина которых равна или больше максимальной толщины скапуса (рис:1/ж/). — Тянь-Шань
M. kryzhanovskii Arnoldi, 1976
19(18) Отстоящие волоски на скапусе явно меньше его максимальной толщины (рис:1/е,з/).
20(21) Усиковые ямки окаймлены явственными морщинками, присоединяющимися к лобным, валикам (рис:3/м/). Несколько крупнее (HL=1,01—1,18; TL=1,37—1,55 м). — Тянь-Шань, Алай
M. juglandeti Arnoldi, 1976
21(20) Морщинки вокруг усиковых ямок нерезкие, часто не доходят до лобных валиков (рис:3/о/). Несколько мельче (HL=0,95—1,02; TL=1,33—1,48 мм). — Ферганская долина, Чаткальский хребет
M. ferganensis Arnoldi, 1976
22(9) Скапус согнут круче, либо без уголка на сгибе, либо угловатый, с вертикальным зубчиком или лопастью (рис:1/л—я, d—f/). Самцы: скапус короче (SI=1,17—1,45 у видов группы lobicornis, либо больше 2 у видов группы schencki).
23(42) Скапус на сгибе без вертикальной лопасти или зубца: либо согнут круто, но без уголка, либо угловатый и лишь с острым кантиком вдоль своей вертикальной части (рис:1/л—у/).
24(25) Грудь в области мезонотума и проподеума сильно уплощена с боков, ее спинная поверхность имеет вид узкого продольного гребня, ограниченного по бокам острыми кантиками, непосредственно переходящими на шипы (рис:4/а/). Скапус на сгибе без уголка (рис:1/л/). Лобные валики искривлены очень слабо, лоб широкий (FI=1,07—1,11; FLI=2,25—2,37). — Южная Сибирь
M. commarginata Ruzsky, 1905
25(24) Грудь не уплощена, обычной формы (рис:4/б/).
26(29) Грудь с очень грубыми, резкими продольными морщинками. Передняя поверхность петиолюса прямая или очень слабо вогнутая, образует с верхней поверхностью прямой или тупой угол, вершинная площадка явственная и не скошена назад (рис:4/в,г/). Скапус на сгибе без уголка. Шипы проподеума длинные, лишь немного короче его горизонтальной поверхности или даже длиннее ее. Лоб относительно широкий (FI=1,06—1,19; FLI=2,22—2,66). Крупнее: HL=0,95—1,33; TL=1,40—1,99 мм.
27(28) Петиолюс с боков с такими же грубыми морщинками, как и на груди, его передняя поверхность очень крутая, образует с вершинной площадкой прямой угол; шипы проподеума немного короче его горизонтальной поверхности (рис:4/в/). — Транспалеаркт
M. sulcinodis Nylander, 1846
28(27) Петиолюс с боков с зернистой скульптурой и нерезкими морщинками, менее грубыми, чем на боках груди, его передняя поверхность образует с вершинной площадкой тупой угол; шипы проподеума мощные, равны его горизонтальной поверхности или даже немного длиннее ее (рис:4/г/). — Приморье, юг Хабаровского края
M. aspersa Kupianskaya, 1990
29(26) Набор признаков иной. Мельче (HL не более 1,15; TL не более 1,64 мм).
30(31) Лоб широкий (FI=1,07—1,15; FLI=2,28—2,55), скапус на сгибе не угловатый. Петиолюс высокий, с явно выраженной передней цилиндрической частью, его передняя поверхность вогнутая, не образует угла с верхней поверхностью; вершинная площадка неявственная, выпуклая, широко закругленная. Шипы проподеума прямые, средней длины, расширенные у основания (рис:4/д/). — Сибирь от Алтая до Тихого океана
M. transsibirica Radtschenko, 1994
31(30) Лоб уже (FI=1,16—1,33; FLI=2,35—2,71). Петиолюс угловатый или заостренный, если нет, то скульптура тела более сглаженная, шипы проподеума короткие и тело явственно двуцветное.
32(37) Скапус согнут резко, с уголком или острым кантиком на своей вертикальной части (рис:1/р,с,у/); если у некоторых особей уголок не выражен, то петиолюс с сильно скошенной назад вершинной площадкой, резко угловатый в профиль (рис:4/з/).
33(34) Петиолюс в профиль не угловатый, его передняя поверхность слегка вогнута и переходит в верхнюю под закругленным углом; вершинная площадка нерезкая, выпуклая, слабо скошена назад. Шипы проподеума средних размеров, прямые, расширенные у основания (рис:4/ж,з/). Голова с негрубыми прямыми продольными морщинками, петлеобразная скульптура развита лишь по бокам головы. — Камчатка, Магаданская обл., север Хабаровского края
M. kamtschatica Kupianskaya, 1986
34(33) Петиолюс в профиль резко угловатый или треугольный (рис:4/ж,з/). Прямые продольные морщинки на голове развиты главным образом на лбу; на темени, висках и затылке преобладает петлеобразная скульптура.
35(36) Петиолюс в профиль треугольный, узко закруглен сверху, вершинная площадка не выражена. Шипы проподеума тонкие, слабо расширены у основания, прямые (рис:4/ж/). Лоб несколько уже (FI=1,19—1,30; FLI=2,50—2,87). — Горы Средней Азии
M. saposhnikovi Ruzsky, 1905
36(35) Петиолюс резко угловатый, его передняя поверхность образует с явственной, сильно скошенной назад вершинной площадкой острый угол. Шипы проподеума расширены у основания, торчащие вверх, часто загнуты внутрь (рис:4/з/). Лоб несколько шире (FI=1,16—1,23; FLI=2,39— 2,70). — Сибирь от Алтая до Тихого океана, северная Монголия
M. angulinodis Ruzsky, 1905
37(32) Скапус на сгибе без уголка и без острого кантика на своей вертикальной части (рис:1/о,и/).
38(39) Петиолюс с прямой передней поверхностью, вершинная площадка сильно скошена назад, так что узелок в профиль клиновидный. — Корея
M. incurvata Collingwood, 1976
39(38) Петиолюс иной формы.
40(41) Лоб шире (FI=1,18—1,31; FLI=2,35—2,46). Лоб, темя и затылок с немногочисленными грубыми продольными морщинками, их число на уровне верхнего края глаз не более 10—12; между глазами и теменем развиты грубые петли. Грудь и петиолюс сбоку с грубыми продольными морщинками. Грудь низкая, мезопроподеальное вдавление очень слабое. Петиолюс высокий, его передняя поверхность вогнутая, образует с верхней поверхностью нерезкий уголок; верхняя поверхность дугообразно переходит в заднюю; шипы проподеума короткие, тонкие, слабо расширены у основания, слегка загнуты концами вниз и направлены более или менее назад (рис:4/и/). Тело одноцветное, оранжево-бурое. Самцы: скапус и особенно голени с короткими отстоящими волосками (рис:2/м,м'/). — Южная Сибирь, северная Монголия
M. kasczenkoi Ruzsky, 1905
41(40) Лоб уже (FI=1,21—1,26; FLI=2,48—2,67), но лобные валики слабо изогнуты и лобные лопасти расставлены нешироко (вследствие чего низкие показатели FI). Лоб и темя с негрубыми многочисленными продольными морщинками, их число на уровне верхнего края глаз больше 15; на боках головы негрубая петлеобразная скульптура. Морщинки на боках груди негрубые, извилистые, часто петлеобразные. Скульптура петиолюса сглаженная, шагреневая, с негрубыми штриховатыми морщинками. Мезопроподеальное вдавление явственное, часто резкое. Передняя поверхность петиолюса вогнутая, его узелок в профиль в виде широко закругленного на вершине равнобедренного треугольника (рис:4/к/). Тело ярко двуцветное. Самцы: скапус и голени с длинными отстоящими волосками (рис:2/л,л'/). — Камчатка, Магаданская обл., Хабаровский край
M. bicolor Kupianskaya, 1986
42(23) Скапус согнут резко, с вертикальной лопастью или хотя бы с небольшим зубцом (рис:1/ф—я,d/).
43(50) Лоб очень узкий (FI=1,35—2,74; FLI=2,92—7,38). Самцы: скапус короткий (SI больше 2).
44(45) Лобные валики очень сильно искривлены, лоб самый узкий из всех известных видов рода (FI=2,13—2,77; FLI=5,56—7,38); лопасть на сгибе скапуса очень массивная (рис:1/d/). Самцы: скапус относительно длинный (SI=2,48—2,76), с длинными отстоящими волосками (рис:2/з/). — Балканы, Турция, Закавказье
M. ravasinii Finzi, 1926
45(44) Лобные валики искривлены слабее, лоб шире (FI не больше 1,95, FLI не больше 4,9); скапус на сгибе с некрупной лопастью или зубцом (рис:1/з—я/).
46(47) Лобные валики изогнуты слабее, лоб шире (FI=1,35—1,46; FLI=2,92—3,50); скапус с зубцом, без развитой лопасти (рис:1/ю/). Мезопроподеальное вдавление слабое, часто отсутствует; грудь с боков с грубыми прямыми продольными морщинками; шипы проподеума короткие, прямые, слабо расширенные у основания (рис:5/е/). Самцы: скапус короткий, слабо изогнутый (SI=3,14—3,61), скапус и ноги с короткими отстоящими волосками (рис:2/ш,ш'/). — Степи и юг лесостепей от Чехословакии до Алтая, Закавказье, западный Копетдаг, Тянь-Шань
M. lacustris Ruzsky, 1905
47(46) Лобные валики изогнуты сильнее, лоб уже (FI=1,47—1,94); скапус с лопастью или зубцом. Бока груди с негрубыми, часто волнистыми, прерывистыми морщинками; шипы проподеума длинные, часто изогнутые, расширенные у основания (рис:5/з/). Самцы: скапус длиннее (SI не более 2,9) (рис:2/ч,ш/).
48(49) Лоб шире (FI=1,47—1,57, FLI=3,76—4,0), скапус на сгибе с зубчиком (рис:1/я/). Шипы проподеума тоньше, загнуты книзу; мезопроподеальное вдавление слабое. Самцы: скапус и ноги с длинными отстоящими волосками, петиолюс короткий и высокий, массивный (рис:2/щ,щ'/; рис:5/ж/). — Южное Закавказье
M. caucasicola Arnoldi, 1934
49(48) Лоб уже (FI=1,53—1,94; FLI=3,61—4,88), скапус на сгибе с лопастью (рис:1/э/). Шипы проподеума расширены у основания, длинные; мезопроподеальное вдавление резкое. Самцы: скапус и ноги с короткими отстоящими волосками, петиолюс не выше постпетиолюса, не массивный (рис:2/ч,ч'/; рис:5/и/). — Транспалеаркт
M. schencki Emery, 1895
50(43) Лоб шире, если такой же узкий (FI=1,43—1,62; FLI=3,05—4,10), то петиолюс с очень короткой передней цилиндрической частью и крутой передней поверхностью, образующей с верхней поверхностью острый угол (рис:5/а/). Самцы: скапус сравнительно длинный (SI меньше 1,5).
51(52) Скапус на сгибе с большой, направленной косо вперед лопастью (рис:1/щ/). — Юго-восток Алтая
M. zojae Radtschenko, 1994
52(51) Скапус на сгибе с зубчиком или с направленной вверх лопастью (рис:1/ф—ш/).
53(54) Лоб узкий (FI=1,43—1,62; FLI=3,05—4,10). Петиолюс с короткой передней цилиндрической частью, его передняя поверхность крутая, вогнутая и образует с верхней резкий, часто острый угол. Вершинная площадка явственная, несколько скошенная назад (рис:5/а/). Лопасть на сгибе скапуса обычно крупная, но варьирует до небольшого зубчика, что более характерно для горных популяций. — Бореальный вид
M. lobicornis Nylander, 1846
54(53) Лоб шире (FI=1,24—1,45; FLI=2,48—2,95), форма петиолюса иная. Скапус с зубчиком или с небольшой лопастью.
55(58) Петиолюс высокий, с почти прямой передней поверхностью, в профиль треугольный, с заостренной вершиной, вершинная площадка не выражена (рис:5/б,в/).
56(57) Шипы проподеума длинные, толстые, в профиль направлены вверх и загнуты внутрь (см. сверху) (рис:4/б/; рис:5/б/). Самцы: скапус у основания сильно изогнут (рис:2/с/). — Сибирь от Алтая до Тихого океана, северная Монголия
M. forcipata Karawajew, 1931
57(56) Шипы проподеума заостренные на концах, прямые, не загибаются внутрь (рис:5/в/). Самцы: скапус слабо изогнут у основания (рис:2/ф/). — Тянь-Шань
M. kirgisorum Arnoldi, 1976
58(55) Петиолюс в профиль не треугольный: либо широко закругленный на вершине, либо низкий, массивный, с нерезкой вершинной площадкой, плавно переходящей в заднюю поверхность (рис:5/г,д/).
59(60) Петиолюс высокий, его передняя поверхность не крутая, вогнутая, вершина широко закругленная, выпуклая (рис:5/г/). Шипы проподеума длинные, острые, расширенные у основания. Мезопроподеальное вдавление глубокое. — Юго-восточное Забайкалье, Амурская обл., Приморье, Япония
M. jessensis Forel, 1901
60(59) Петиолюс низкий, массивный, его передняя поверхность слабо вогнута и образует с верхней нерезкий угол. Вершинная площадка нерезкая, не скошена назад, обычно плавно переходит в выпуклую заднюю поверхность; шипы проподеума тонкие, не расширенные у основания, слегка загнуты вниз и направлены назад (рис:5/д/). Мезопроподеальное вдавление слабое, иногда почти отсутствует. — Южная Сибирь, северная Монголия
M. pisarskii Radtschenko, 1994
61(6) Усиковые ямки не окаймлены дуговидными морщинками (рис:6/а,б/); скапус согнут круто, либо без уголка, либо с горизонтальной закраинкой, зубцом или лопастью (рис:1/f—t/). Самцы: скапус короткий (SI больше 2, лишь у M. sabuleti SI=1,5—1,97).
62(71) Скапус согнут плавно, без уголка на сгибе (рис:1/f—j/).
63(66) Лобные валики искривлены сильнее, лоб уже (FI=1,17—1,56; FLI=2,25—2,65). Тело желтое, слабо скульптированное. — Горы Средней Азии.
64(65) Шипы проподеума очень длинные, палочковидные, на концах заостренные (рис:2/р/). Самцы: скапус очень короткий (SI=2,9—3,0). — Горы Средней Азии
M. orthostyla Arnoldi, 1976
65(64) Шипы проподеума короче, слегка расширенные у основания (рис:2/п/). Самцы: скапус несколько длиннее (SI=2,52—2,76). — Горы Средней Азии, тугаи Амударьи и Сырдарьи
M. tenuispina Forel, 1904
66(63) Лобные валики искривлены слабо, лоб шире (FI не больше 1,14; FLI не больше 2,18). Тело от охристо-оранжевого до темно-бурого. — Европа, Кавказ, юг Западной Сибири (в горах Средней Азии только M. bergi).
67(68) Нижняя часть лба с продольными морщинками, на остальных частях головы развита петлеобразная скульптура (рис:6/а/). Мельче (HL=0,91—1,18; TL=1,29—1,47 мм). Самцы: скапус короткий (SI=2,28— 2,70), 2-й членик жгутика короткий, лишь немного длиннее 3-го (рис:2/d/) — Европа, юг Западной Сибири
M. rugulosa Nylander, 1849
68 (67) Большая часть головы покрыта продольными морщинками, петли развиты лишь между теменем и глазами (рис:6/б/). Самцы: скапус длиннее, если такой же короткий, то 2-й членик жгутика длинный, не менее, чем в 1,5 раза длиннее 3-го (рис:2/g/)
69(70) Шипы проподеума короткие, постпетиолюс низкий, со слабо выпуклой вершиной (рис:6/в/). Самцы: скапус относительно длинный (SI=1,69— 1,97). 2-й членик жгутика короткий, лишь немного длиннее 3-го (рис:2/f/). — Степи и лесостепи от низовьев Днепра до Якутии, горы Средней Азии, северная Монголия
M. bergi Ruzsky, 1902
70(69) Шипы проподеума длинные, постпетиолюс высокий, его вершина сдвинута назад (рис:6/г/). Самцы: скапус короткий (SI=2,44—2,75), 2-й членик жгутика длинный, не менее, чем в 1,5 раза длиннее 3-го (рис:2/g/). — Европа, юг Западной Сибири, Северный Кавказ
M. gallienii Bondroit, 1919
71(62) Скапус согнут круто, на сгибе с уголком и с продольным зубчиком, закраинкой или лопастью (рис:1/l—t/).
72(75) Скапус без развитой лопасти, согнут под тупым углом, лишь с закраинкой или зубцом (рис:1/l—m/).
73(74) Петиолюс в профиль закругленно-треугольный, очень редко с нечеткой, короткой вершинной площадкой; шипы проподеума тонкие, не расширенные у основания; скульптура груди и стебелька из грубых правильных продольных морщинок (рис:6/д/). Тело темное, красновато-бурое. Самцы: отстоящие волоски на голенях не больше максимальной толщины голеней (рис:2/h/), скапус короче (SI=3,17—3,50). — Степи от низовий Днепра до восточного Казахстана
M. stangeana Ruzsky, 1902
74(73) Петиолюс с нерезкой скошенной назад вершинной площадкой; шипы проподеума длинные, расширенные у основания; грудь и стебелек сбоку с продольными, слегка волнистыми морщинками (рис:6/е/). Тело охристо-рыжее различных оттенков. Самцы: отстоящие волоски на голенях явно больше максимальной толщины голеней (рис:2/i/), скапус длиннее (SI=2,03—2,57). — Чехословакия, Болгария, Молдова, Крым, Кавказ, Закавказье, Копетдаг
M. bessarabica Nassonov, 1889
75(72) Скапус с лопастью, согнут под прямым углом (рис:1/n—t/).
76(77) Петиолюс без четкой вершинной площадки (рис:6/ж/). Лопасть скапуса всегда выражена, часто массивная (рис:1/t/). Лобные валики сильно искривлены, лобные лопасти широкие, слегка приподняты над уровнем лба, лоб очень узкий (FI=1,70—2,06; FLI=3,29—3,94). Самцы: скапус короткий (FI(?SI)=2,65—2,90). — Германия, Чехословакия, Венгрия, Болгария, степи и лесостепи Русской равнины, северный Казахстан
M. slovaca Sadil, 1951
77(76) Петиолюс с резкой, явственной горизонтальной площадкой (рис:6/з/), если площадка нерезкая, скошенная назад, то лоб шире (FI меньше 1,7; FLI меньше 3,05).
78(79) Лопасть на сгибе скапуса массивная (рис:1/s/), лоб уже (FI=1,59—1,86; FLI=3,04—3,50). Самцы: скапус относительно длинный, равен 4—4,5 членикам жгутиков антенн (SI=1,50—1,97), с короткими отстоящими волосками (рис:2/l/). — Европа, юг Западной Сибири
M. sabuleti Meinert, 1860
79(78) Лопасть на сгибе скапуса небольшая, иногда зубцевидная (рис:1/n/), лоб шире (FI=1,41—1,68; FLI=2,60—3,04). Самцы: скапус короткий, равен 3—3,5 членикам жгутиков антенн (SI=2,29—2,95), с длинными отстоящими волосками (рис:2/j/). — Транспалеаркт
M. scabrinodis Nylander, 1846
80(5) Боковые края клипеуса килевидно приподняты, киль резко окаймляет спереди усиковую впадину (рис:6/и/) (как у видов рода Tetramorium).
81(82) Скапус на сгибе толстый, не тоньше, чем на середине своей длины, с лопастевидными выступами (рис:1/w,w'/); крупнее: HL=1,11—1,15; TL=1,65—1,74 мм. Самцы: членики стебелька матовые. — Приморье
M. excelsa Kupianskaya, 1990
82(81) Скапус на сгибе явно тоньше, чем на середине своей длины без лопастевидных выступов у основания (рис:1/v/); мельче: HL=0,98—1,08; TL=1,44—1,54 мм. Самцы: членики стебелька блестящие. — Приморье, юг Хабаровского края, Амурская обл.
M. carinata Kupianskaya, 1990.
СПИСОК ЛИТЕРАТУРЫ
Арнольди К. В., 1970. Обзор муравьев рода Myrmica (Hymenoptera, Formicidae) европейской части СССР//Зоол. журн. Т. 49. Вып. 12. С. 1829—1843.— 1976. Муравьи рода Myrmica Latr. Средней Азии и Южного Казахстана//Там же. Т. 55. Вып. 4. С. 547—558.
Арнольди К. В., Длусский Г. М., 1978. Семейство Formicidae — муравьи//Определитель насекомых европейской части СССР. М.: Наука. Т. 3. Ч. 1. С. 519—556.
Длусский Г. М., Союнов О. С, Забелин С. И., 1990. Муравьи Туркменистана. Ашхабад: Ылым. С. 1—273.
Купянская А. Н., 1990. Муравьи Дальнего Востока СССР. Владивосток: ДВО АН СССР. С. 1—258.
Рузский М. Д., 1905. Муравьи России. Казань. С. 1—798.
Тарбинский С., 1976. Муравьи Киргизии. Фрунзе, Илим. С. 1—217.
Agosti D., Collingwood C. A., 1987. A provisional list of the Balkan ants (Hymenoptera, Formicidae) and a key of the worker caste. I. Synopsis list. II. Key of the worker caste, including the European species without the Iberian//Mitt. Schweiz. Entomol. Ges. Bd. 60. H. 1—2, 3—4. S. 51—62, 261—293.
Seifert В., 1988. A taxonomic revision of the Myrmica species of Europe, Asia Minor, and Caucasus (Hymenoptera, Formicidae)//Abh. Ber. Naturkind. Bd. 62. H. 3. S. 1—75.
Институт зоологии АН Украины, Киев
Поступила в редакцию 9 декабря 1993 г.
A. G. RADCHENKO
A KEY ТО THE SPECIES OF THE GENUS MYRMICA (HYMENOPTERA, FORMICIDAE) OF THE CENTRAL AND EASTERN PALAEARCTIC
Institute of Zoology, Ukrainian Academy of Sciences, Kiev, Ukraine
Summary
A key to 42 species of Myrmica from Central and Eastern Palaearctic is given.
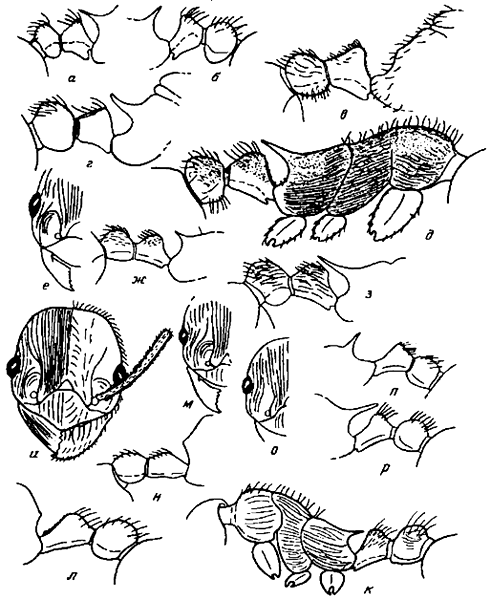
Рис. 3. Стебелек и проподеум рабочих в профиль (а, б, ж, з, п, р), то же самое (в, г) грудь и стебелек рабочих в профиль (д, к) голова рабочего спереди (и), часть головы рабочих с усиковой ямкой (е, м, о): а — M. arnoldii (паратип); б — M. tschekanovskii (паратип); в — M. luteola (паратип); г — M. forcipata; д — M. kurokii; е, ж —

Рис. 1. Скапус рабочих (a—я', d—t — сбоку; n'—t' — сверху): а — M. arnoldii (голотип); б — M. tschekanovskii (паратип); в —
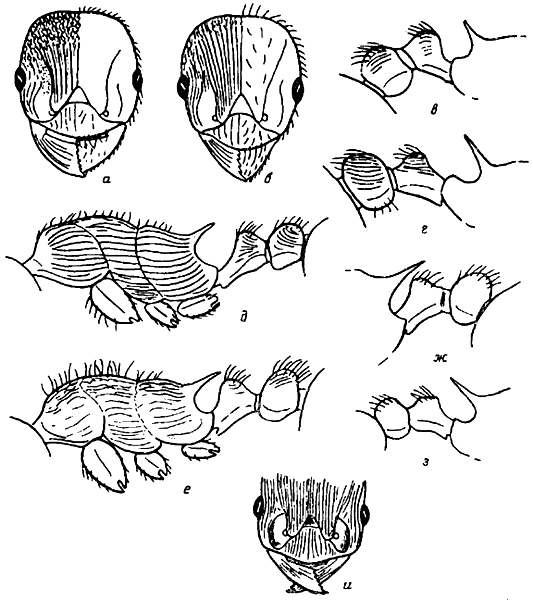
Рис. 6. Голова рабочих спереди (а, б, и), стебелек и проподеум в профиль (в, г, ж, з), грудь и стебелек в профиль (д, е): а — M. rugulosa; б, в — M. bergi (синтип); г — M. gallienii; д — M. stangeana (лектотип): е — M. bessarabica; ж — M. slovaca (паратип); з — M. scabrinodis; и — M. carinata (голотип)

Рис. 2. Скапус (а—я, d—m — сбоку) и задняя голень а'—я', d'—m' самцов: а — M. arnoldii, б — M. tschekanovskii (голотип), в —
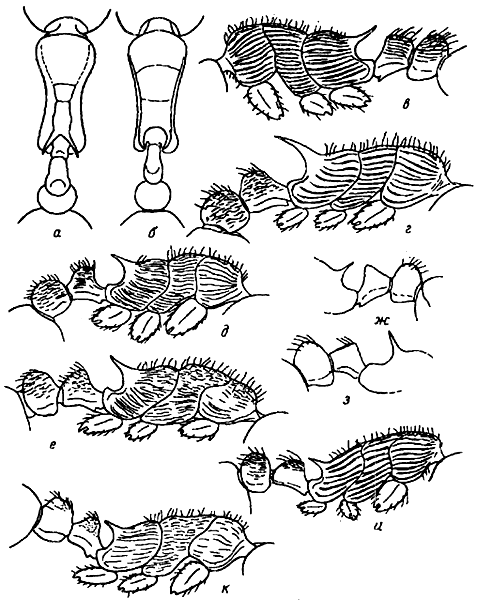
Рис. 4. Грудь рабочих сверху (а, б) грудь и стебелек в профиль (в—е, и, к), стебелек и проподеум в профиль (ж, з): а — M. commarginata (неотип), б — M. forcipata (синтип), в — M. sulcinodis, г — M. aspersa (голотип), д — M. transsibirica (голотип), е — M. kamtschatica (паратип), ж — M. saposhnikovi (неотип), з — M. angulinodis (неотип), и — M. kasczenkoi (неотип), к — M. bicolor (паратип)
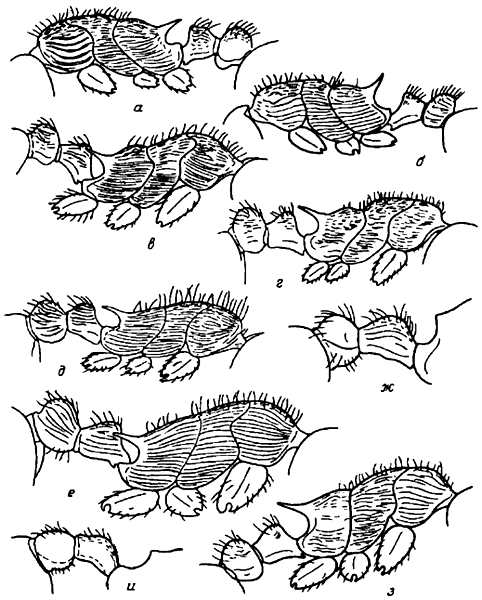
Рис. 5. Грудь и стебелек рабочих в профиль (а—е, з), стебелек самцов в профиль (ж, и); а — M. lobicornis; б — M. forcipata (синтип); в — M. kirgisorum (паратип); г — M. jessensis; д — M. pisarskii (голотип); е — M. lacustris; ж — M. caucasicola; з, и — M. schencki
МУРАВЬИ - семейство общественных насекомых подотряда жалящих перепончатокрылых. Образуют сложные семьи, включающие несколько каст, выполняющих различные функции (фуражиров, солдат и др.). Длина рабочих муравьев 0,8-30 мм, самки крупнее. Ок. 10 тыс. видов. Более разнообразны и многочисленны в тропиках, северная граница распространения лесотундра. Гнезда в почве, древесине, иногда надземные сооружения (муравейники); существуют и бродячие виды, не имеющие постоянного гнезда. Уничтожают насекомых-вредителей, участвуют в почвообразовании. Во многих странах ряд видов муравьев и их гнезд охраняются.
Всего 141 вид.
эволюционно сложившаяся, пространственно ограниченная, длительно самоподдерживающаяся, однородная экологическая система, в которой функционально взаимосвязаны живые организмы и окружающая их абиотическая среда.
первый членик усика, обычно намного длиннее члеников жгутика
 Myrmica rubra (Linnaeus, 1758) — рыжая мирмика
Myrmica rubra (Linnaeus, 1758) — рыжая мирмикаСтарое название: Myrmica deplanata Emery, 1921
Subspecies: M. sulcinodis eximia Kupyanskaya, 1990
Старое название: M. aspersa Kupyanskaya, 1990
Subspecies: Myrmica scabrinodis ussuriensis, Kuznetsov-Ugamsky
Старое название: Myrmica slovaca Sadil, 1952
Старое название: Myrmica limanica Karavaiev, 1934
Старое название: M. carinata Kupyanskaya,1990
особый сегмент брюшка. У муравьев бывает двучлениковый (петиолюс + постпетиолюс) и одночлениковый (петиолюс - чешуйка). Количество члеников и форма стебелька очень важны для идентификации вида. Например у муравьев семейства Formicinae стебелек одночлениковый, а у Myrmicinae - двучлениковый.
часть груди муравья. Состоит из: скутума и скутеллюма. (Извините, информация нуждается в дополнении/уточнении)
совокупность особей одного вида, длительно занимающая определенное пространство и воспроизводящая себя в течение большого числа поколений. В современной биологии популяция рассматривается как элементарная единица процесса эволюции, способная реагировать на изменения среды перестройкой своего генофонда.
Всего 415 видов
класс беспозвоночных типа членистоногих. Тело разделено на голову, грудь и брюшко, 3 пары ног, у большинства крылья. Дышат трахеями. Развитие, как правило, с метаморфозом яйцо, личинка, нимфа (или куколка), взрослое насекомое. Самая многочисленная и разнообразная группа животных на Земле.






