






Разделение функций среди рабочих особей в семьях общественных насекомых является классическим примером реализации адаптивной изменчивости в популяциях. Для муравьев хорошо исследованы ситуации, когда выполняемые функции обеспечиваются морфологической кастовой изменчивостью: специфической формой челюстей, головы, существенными различиями в размерах [ 1 ]. Однако у многих видов морфологических различий между функциональными группами нет, и разделение труда основано только на различиях в поведении [ 2 ]. Относительная роль врожденных и приобретенных в процессе онтогенеза структурно-функциональных параметров разделения труда у муравьев (как и у других общественных насекомых) оставалась невыясненной. Причина неудач, возможно, заключалась в методологическом подходе, основанном на изучении поведения "безличных" функциональных групп в семье (сборщиков пади тлей, охотников, охранников, нянек, строителей). Вместе с тем исследования на индивидуальном уровне позволили выявить в пределах одной из специализированных функциональных групп рыжих лесных муравьев, - сборщиков пади систему "профессионального" разделения труда: пастухов, охранников, разведчиков и транспортировщиков. При этом оказалось, что возраст не является существенным фактором, влияющим на выбор муравьем профессии [ 3 ]. Можно полагать, что у муравьев профессиональная специализация реализуется в процессе индивидуального развития на основе сочетания когнитивных (познавательных) и психофизиологических характеристик, таких, как уровень и количество агрессивных реакций, способность обучаться избегать опасности или решать определенные поисковые задачи.
Для экспериментальной проверки этой гипотезы мы в качестве первого шага исследовали проявление и раннее развитие агрессивного поведения у рыжих лесных муравьев Formica aquilonia Yarrow. В основе схемы экспериментов лежит представление о видотипическом образе врага, разработанное Тинбергеном [ 4 ] на примере птиц. Ранее выявлено, что для рыжих лесных муравьев одним из образов врага служат их конкуренты - хищные жужелицы, в экспериментах успешно заменяемые моделями с характерными признаками. Тестирование представителей разных функциональных групп показало, что охранники нападают на врагов, тогда как сборщики пади избегают контакта с опасными объектами [5]. Это позволило использовать жужелиц как тестовый объект для выявления профессиональных характеристик муравьев. Мы предположили, что охранники рекрутируются из особей, обладающих наиболее явно выраженным видотипическим образом врага (или набором подобных образов), высоким уровнем агрессии и ранним ее проявлением в имагинальном онтогенезе.
Организация и результаты экспериментов.
В экспериментах сравнивали реакции на врагов у муравьев, взятых из двух пар одинаковых по численности лабораторных семей: базовой (естественной) и "наивной", состоящей из особей, вышедших из куколок в лабораторных условиях и выращенных в изоляции. Муравьев тестировали индивидуально на аренах (15 х 15 см) с предъявлением им живого жука или модели, которую передвигали на нитке со скоростью 6-7 см/с. Гипотезы о различиях между реакциями муравьев на разные объекты проверяли с использованием формулы Фишера для сравнения двух выборочных долей вариант: критерия хиквадрат и критерия Вилкоксона [6]. В первой, предварительной серии опытов участвовали четыре группы муравьев (по семь особей): охранники, охотники и сборщики пади из базовой семьи, а также наивные муравьи возраста около двух недель. Охранников отбирали с крышки формикария, пронося над ним иглу и собирая наиболее агрессивно нападающих на нее муравьев. Охотников отбирали по реакции на потенциальную добычу на жилой арене. Для выявления сборщиков пади на арену выставляли ветки осины с колониями тлей. Для 28 муравьев было проведено 140 тестов. В этой серии опытов выяснено, что если охранники и охотники из базовой семьи агрессивно реагируют не только на жука, но и на модель врага, то наивные муравьи существенно чаще реагируют на живого жука (p < 0.05 по формуле Фишера). Это позволяет предположить, что выделение ключевых признаков врага требует определенной достройки [7]. Предварительные оценки показали, что наименее агрессивно по отношению к врагу ведут себя наивные муравьи и сборщики пади, а охранники и охотники демонстрируют сходный и более высокий уровень агрессии.

Рис. 1. Доля особей группы охранников из базовой семьи, демонстрировавших продолжительные укусы в тестах с моделями и жуком.
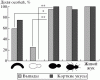
Рис. 2. Доля особей группы муравьев из наивной семьи, демонстрировавших реакцию агрессивных выпадов в тестах с моделями и жуком (критерий хи-квадрат, * p < 0.05, ** p < 0.01).
Во второй серии опытов участвовали две группы по 20 особей, взятые из двух других семей: недифференцированная группа муравьев трех-пятинедельного возраста из наивной семьи и группа охранников из базовой семьи. Муравьев по одному ссаживали с живым жуком в течение 15 мин. Спустя 5-15 мин муравьев тестировали на моделях, предъявляя их в случайном порядке по одной, каждую в течение 15 мин. Всего проведено 200 предъявлений и зафиксированы последовательности и количество поведенческих реакций у 40 особей. Были выявлены следующие реакции на модели и живого жука, ранжированные по увеличению степени агрессивности: 1) раскрытие жвал, 2) выпад с раскрытыми жвалами, 3) короткий укус (менее 5 с), 4) продолжительный укус (более 5 с), 5) "мертвая хватка", 6) преследование объекта агрессии, сопровождавшееся выпадами и короткими укусами; 7) опрокидывание муравья на спину и попытка вцепиться жвалами в конечность жука. Последняя реакция была выявлена только у наивных муравьев и только по отношению к живому врагу. Спектры поведенческих реакций, демонстрируемых муравьями из двух наивных и двух базовых семей на жуков и их модели оказались сходными, что говорит о значительной доли наследственно обусловленной компоненты агрессивного поведения. Отмечены два существенных различия: 1) наивные муравьи никогда не демонстрировали мертвую хватку, тогда как 20% членов базовых семей проявляли эту реакцию; 2) 65% наивных муравьев, не сумев закрепиться жвалами на жертве, демонстрировали падение на спину, а у членов базовых семей эта реакция отсутствовала. Эти различия связаны с формированием мертвой хватки как завершающего акта агрессивного поведения, который, по-видимому, требует достройки за счет накопления опыта.
Сравнение реакций на жужелиц и их модели у представителей разных функциональных групп позволило выявить следующие тенденции. Охранники использовали наиболее агрессивные реакции при взаимодействии с живыми жужелицами. Они также агрессивно реагировали на все модели, в том числе несущие единственный ключевой признак. Наибольшую агрессию вызывали модели, приближенные по своему облику к оригиналу (рис:1). Важно отметить, что у наивных муравьев, так же как и у муравьев из базовых семей, белый цвет модели выключал проявление агрессивных реакций (выпадов и коротких укусов), а несимметричная форма модели снижала их проявление (рис:2; см. также [5]). Это позволяет полагать, что наивные муравьи при распознавании врагов используют те же визуальные признаки, что и взрослые особи.
Сравнение характера поведения на индивидуальном уровне позволило нам, хотя пока на ограниченном материале, предположить, что распределение муравьев по функциональным группам основано на существенных различиях в ранних проявлениях агрессивности. Дело в том, что из 27 наивных муравьев обеих семей три особи демонстрировали и продолжительный укус, и реакцию преследования в тестах с жужелицей. По количеству агрессивных реакций на живого жука эти муравьи сопоставимы с взрослыми охранниками. Можно предположить, что именно из подобных особей, рано проявляющих высокую агрессивность, и рекрутируются будущие охранники и охотники. Вполне возможно, что агрессивные муравьи обладают врожденным запретом на формирование такой естественной реакции, как обучение избегать опасности. Это, в свою очередь, может быть связано с проявлением когнитивной специализации [8]: сочетание облегченного формирования одних ассоциативных связей с запретом на формирование других. Эта гипотеза нуждается в проверке. На данном этапе наши результаты дают возможность полагать, что основную роль в формировании профессиональной специализации у муравьев играет сочетание психофизиологических характеристик. Для охранников и охотников характерны высокий уровень агрессии и рано проявляющиеся реакции на сочетание признаков врага. Способности выделять ключевые признаки и эффективно завершать нападение требуют достройки за счет индивидуального опыта и, возможно, простых форм социального обучения, как это было показано ранее для охотничьего поведения муравьев [9].
Авторы благодарны чл.-кор. РАН В.И. Евсикову за плодотворную дискуссию и ценные замечания.
Исследования поддержаны грантами РФФИ (05-04-48104), Совета по грантам Президента РФ (НШ - 1038.2006.4д) и Президиума РАН по программе "Происхождение и эволюция биосферы".
СПИСОК ЛИТЕРАТУРЫ
- Holdobler В., Wilson, E.O. The Ants. Harvard: The Belknap Press of Harvard Univ. Press, 1990. 780 p.
- Dobrzanska J. // Acta biol. exp. 1958. V. 18. P. 55-67.
- Резникова Ж.И., Новгородова ТА. // ДАН. 1998. Т. 359. < 4. C. 572-574.
- Tinbergen N. The Study of Instinct. Oxford: Clarendon, 1951. 380 p.
- Дорошева Е.А., Резникова Ж.И. // Зоол. журн. 2006. Т. 85. < 2. С. 183-191.
- Урбах В.Ю. Биометрические методы. М.: Наука, 1975. 425 с.
- Лобашев М.Е. В кн.: Исследования по генетике. Л.: ЛГУ, 1961. Т. 1. С. 1-11.
- Reznikova Zh. Animal Intelligence: from Individual to Social Cognition. Cambridge: Univ. Press, 2007. 497 p.
- Резникова Ж.И., Пантелеева С.Н. // ДАН. 2005. Т. 401. < 1. С. 139-144.
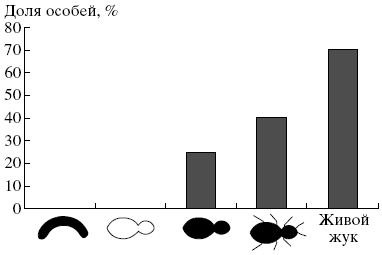
Рис. 1. Доля особей группы охранников из базовой семьи, демонстрировавших продолжительные укусы в тестах с моделями и жуком.
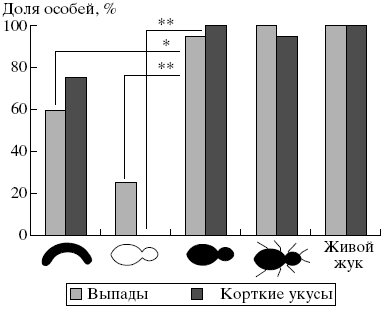
Рис. 2. Доля особей группы муравьев из наивной семьи, демонстрировавших реакцию агрессивных выпадов в тестах с моделями и жуком (критерий хи-квадрат, * p < 0.05, ** p < 0.01).
основная форма существования общественных насекомых. Состоит из репродуктивных (самки, самцы) и функционально бесполых особей (рабочие).
живут колониями (пчелы, муравьи, термиты, некоторые осы). В состав колонии, напр. у термитов, входят половые особи (самцы и матки обеспечивают размножение), солдаты (защиту) и рабочие (собирание корма, строительство гнезда и т. д.). Солдаты и рабочие бесплодны; половые особи и солдаты сами не способны питаться; таким образом, члены колонии полностью зависят друг от друга. Разделение функций и сложные взаимодействия разных особей в колониях общественных насекомых обеспечиваются рядом особенностей в их строении и поведении; последнее регулируется с помощью выделяемых ими химических веществ (феромонов).
совокупность особей одного вида, длительно занимающая определенное пространство и воспроизводящая себя в течение большого числа поколений. В современной биологии популяция рассматривается как элементарная единица процесса эволюции, способная реагировать на изменения среды перестройкой своего генофонда.
у муравьев имеется три основные касты: самцы, самки и рабочие (бесплодные модифицированные самки).
устойчивые группы особей, занятых в семье выполнением определенных функций. Входят в состав полиэтических групп.
(rp. ontos - сущее, существо) - индивидуальное развитие организма с момента зарождения до смерти.
сладкая густая жидкость, выделяемая тлями, червецами и другими насекомыми, питающимися соком растений. Медоносная пчела собирает падь с листьев и перерабатывает в падевый мед.
подотряд насекомых отряда равнокрылых. Длина 0,5-6 мм. Св. 2,5 тыс. видов. Распространены главным образом в умеренном поясе Северного полушария. Живут колониями на растениях. Многие тлевые опасные вредители, некоторые переносчики ряда вирусных заболеваний растений. Сахаристые испражнения тлевых (падь) привлекают муравьев, которые охраняют тлевых.
искусственный муравейник.
(лат. mandibule — челюсть, от mando — жую, грызу), первая пара челюстей у ракообразных, многоножек и насекомых. Синоним: жвала.
необратимое историческое развитие живой природы. Определяется изменчивостью, наследственностью и естественным отбором организмов. Сопровождается приспособлением их к условиям существования, образованием и вымиранием видов, преобразованием биогеоценозов и биосферы в целом.






